HINT 23A: Recall that the mutation load depends only on U) (see p. 552).
HINT 23B: Think about an allele on one of the chromosomes, which changes the genome-wide mutation rate. Will such an allele increase?
HINT 23C: Assuming that the effects of deleterious alleles on fitness multiply together, the number of deleterious alleles will follow a Poisson distribution; see Box 28.5).
HINT 23D: Begin by calculating the expected number of mutation-free offspring.
HINT 23E: Ignore the initial effects of random drift, which will cause most mutations to be lost, even if they are favorable. In the absence of deleterious mutations, the probability of fixation would be approximately 2s (depending on the distribution of offspring number [see p. 490], but the presence of mutations complicates this).
HINT 23F: Consider what will happen both if the favorable allele has smaller effects and if it has larger effects.
HINT 23G: A qualitative answer will do.
HINT 23H: In a stable outcrossing population, each plant produces on average one seed itself and fertilizes one seed on another plant. Track the numbers of heterozygotes and homozygotes for the selfing allele from one generation to the next.
HINT 23I: Either find the leading eigenvalue, or follow the numbers through several generations to find the eventual rate of increase.
HINT 23J: Work out how many generations it will take for allele frequencies to be equal (see pp. 468–469 and Box 17.1).
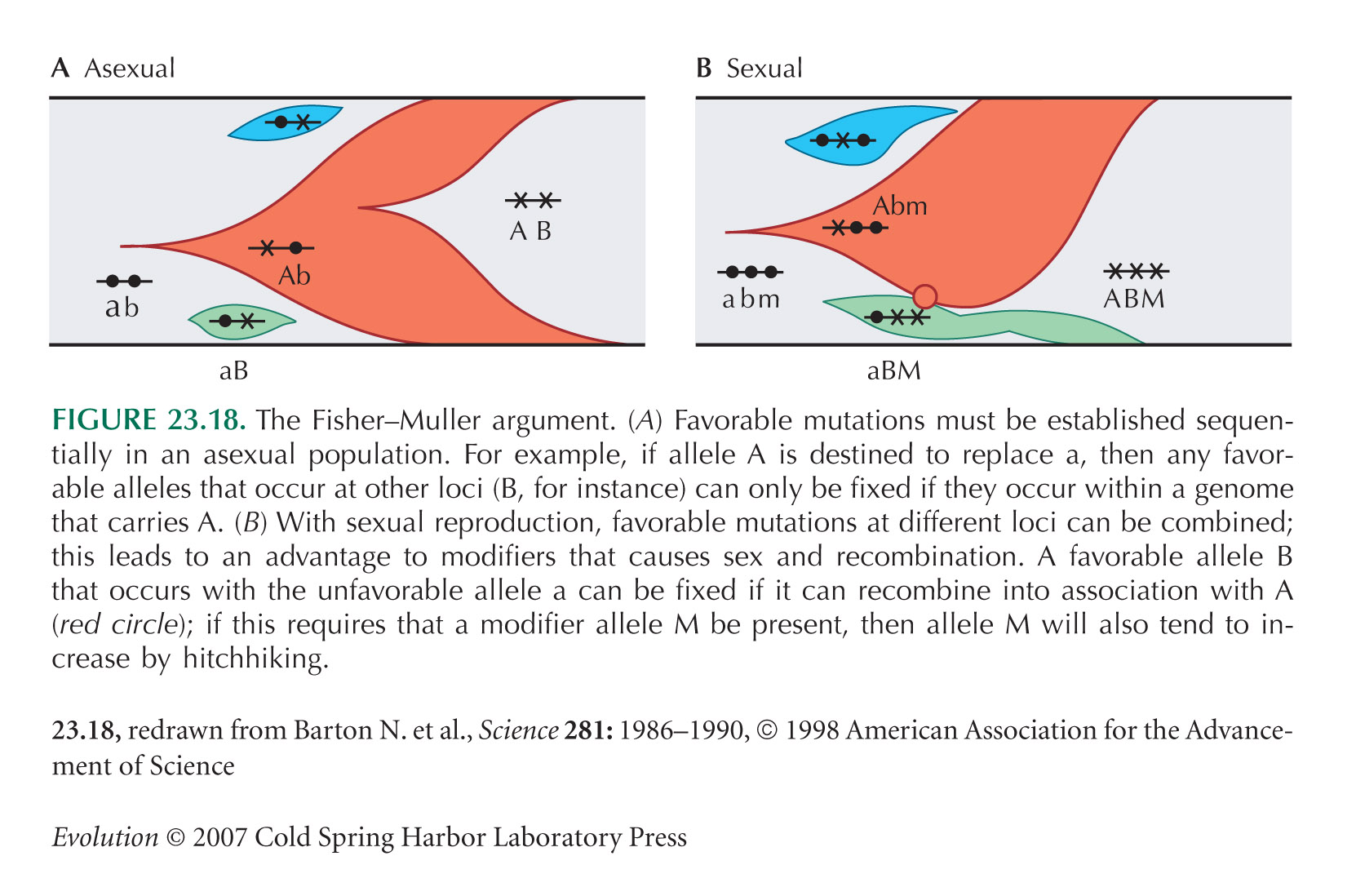
HINT 23K: An approximate answer will be fine; see also Figure 23.18.
HINT 23L: Think about the rate of substitution for both sexual and asexual populations.
HINT 23M: Think about the chance that one of the parents carries the rare allele, and how this will be passed on to offspring given intense competition.
HINT 23N: Begin by working out the relative survival of each genotype in each environment.
HINT 23O: Write the frequencies of the four genotypes at the start of the generation as {1/4 + D, 1/4 – D, 1/4 – D, 1/4 + D}, where D is the strength of linkage disequilibrium (p. 433); then, find how these change as a result of selection within patches. Recall that random mating halves the linkage disequilibrium between unlinked loci (p. 434).
HINT 23P: Compare the mean fitness when all four genotypes are present with the mean fitness if only the two fittest genotypes are present.
HINT 23Q: Using a two-generation cycle (which is equivalent to one alternating cycle), think about the effect of selection and random mating upon genotype frequencies.
HINT 23R: The variance of the average of the two parents is half the variance of each parent.
HINT 23S: Because exp(–x) ~1 – x for small x, the fitness can be approximated by 1 – z2/2Vs. Therefore, we need just the average of this formula.
HINT 23T: Multiply the initial distribution by the relative fitness to get the probability of a trait value after selection.
HINT 23U: It may be easier to assume a specific strength of selection, say Vs = 10Vf.
HINT 23V: A qualitative answer will do.
HINT 23W: With unlinked loci, linkage disequilibrium halves in each generation.
HINT 23X: Assume that δ is small.
HINT 23Y: Recall page 552.
HINT 23Z: This will approximate to a Poisson distribution.
HINT 23AA: Assume that in a stable population, both wild type and haplodiploids produce one son and one daughter. Assuming that allele A is rare, you must follow the numbers of Aa females, of diploid Aa males, and haploid A males. These numbers can be iterated through time for a numerical example; better, the methods of Chapter 28 can be used to find the long-term rate of increase.
HINT 23BB: Assume that the mutation rate at each locus is very low, so that lethal homozygotes are negligibly rare in diploids.
HINT 23CC: You can ignore the weak selection s against recessive lethals as heterozygotes, because they will mainly be eliminated when exposed in males.
HINT 23DD: Calculate the expected number of recessive lethals (inherited + new) for daughters and sons.
HINT 23EE: Assume that the population is fixed for maximum gamete size M.
HINT 23FF: Work out the net fitness for each gamete type; at equilibrium, these must be equal. For further detail and references, see Web Notes.
HINT 23GG: We can measure E and 1/β relative to the substrate concentration, X.
HINT 23HH: Sketch a graph of fitness against enzyme concentration.
{kind=link}