Trade-offs May be Revealed by Patterns of Genetic Variation or by the Response to Artificial Selection
As we have seen, optimality arguments must depend on constraints on what is possible. There are trade-offs between different components of fitness, such that not all components can be maximized at the same time. One might therefore expect trade-offs to be reflected in negative correlations between fitness components. For example, if an increase in early reproduction is made at the expense of later survival, then one would expect that within a population, individuals that reproduce early will tend to die early, and vice versa. Moreover, one would expect negative correlations to be seen at the genetic as well as the phenotypic level. Individuals that produce offspring with earlier reproduction will also tend to have offspring that live less long. Such genetic correlations are especially important, because, as we saw on page 480, they determine the immediate direction of change caused by selection. Thus, in the selection experiments on “young” versus “old” lines discussed above (Box 20.2), a shift to earlier reproduction in “young” lines was accompanied by a decrease in life span; the opposite pattern was seen in the “old” lines. These negative correlations in the response to selection of different parts of the life history reflect negative genetic correlations in the base population.
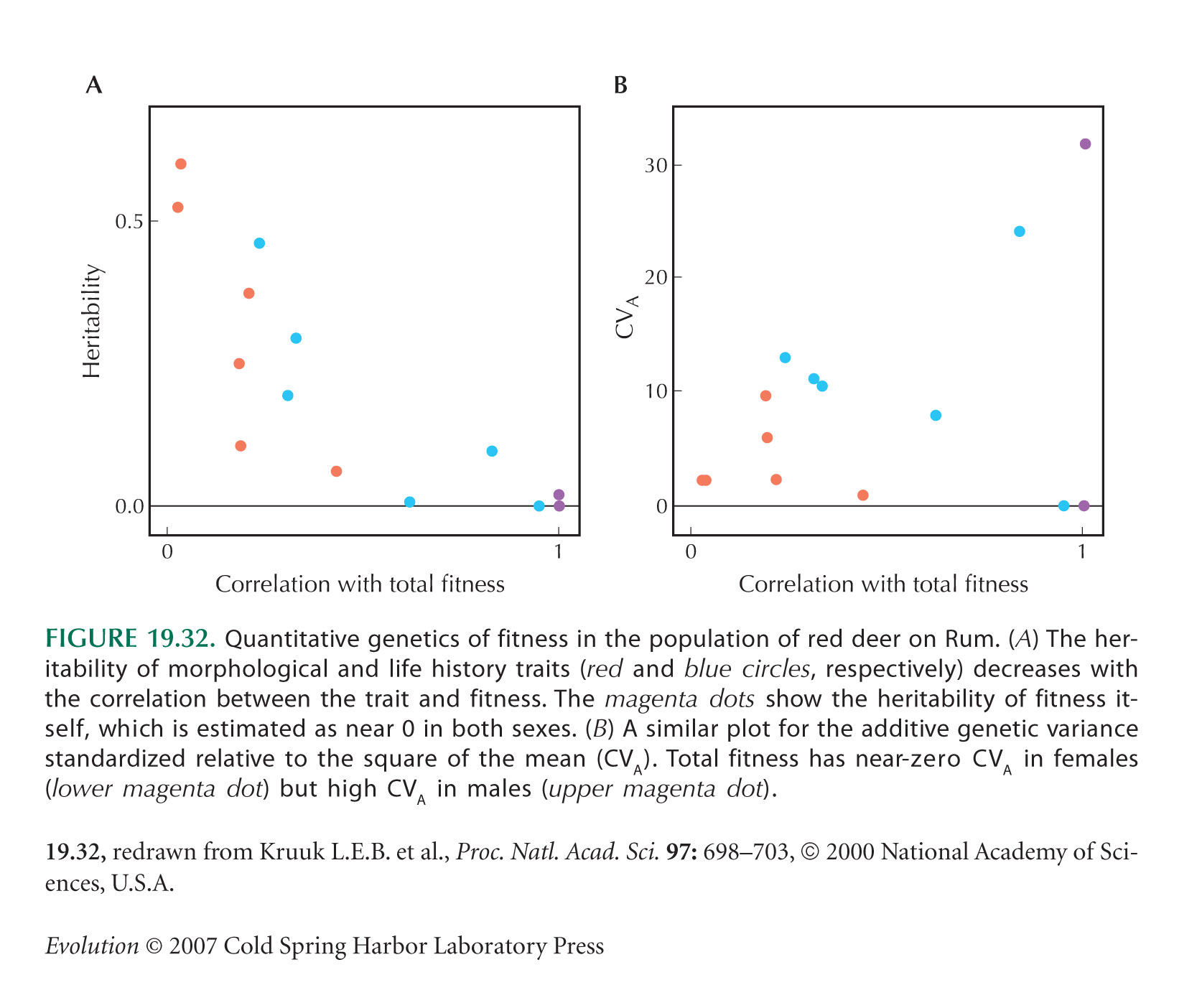
Considerable effort has been put into measuring genetic correlations for a variety of traits. In the , we saw that in order to find the causes of fitness variation, that is, to find which traits are under direct selection, the pattern of genetic correlations must be measured. Studies of genetic correlation have also been motivated by the idea that if trade-offs between different aspects of the life history can be demonstrated, then the actual life history can be understood as a compromise between these different aspects. For example, senescence may evolve because there is a trade-off between performance in early life and late life, in which case we might expect to see negative genetic correlations in the life history: Genotypes that give high fitness in youth should age faster. Often, however, positive correlations are found at both phenotypic and genetic levels. Extensive work on life-history variation within laboratory populations of Drosophila has shown a mixture of negative and positive correlations between early and late life performance. In red deer, the additive genetic variance of traits is higher for traits more closely associated with fitness, rather than lower (e.g., Fig. 19.32B). Yet, if fitness components were negatively correlated as a result of trade-offs, we would expect the opposite pattern, leading to low variance in net fitness. Such observations are surprising, because they suggest that selection could increase several different components of fitness at the same time. How can we explain the unexpected prevalence of positive genetic correlations between fitness components?
There are several reasons why positive correlations might be seen, even when the different traits are tightly constrained. First, not all the relevant components are measured. If many different traits are correlated, each individual correlation will be small (~1/n if there are n traits), and many may be positive rather than negative. Overall, the whole pattern of correlation is constrained, and correlations may on average be negative. However, this pattern may be hard to discern. In practice, the problem is much worse, because not all fitness components can be measured, and none can be estimated accurately (pp. 525–530).
A more fundamental explanation is that variation within populations need not reflect the boundaries on what is possible. Variation in environment—amount of food, disease, and so on—will tend to affect many components of fitness in the same way and so produce positive correlations (recall the red deer example of Fig. 19.8). Similarly, deleterious mutations may also reduce many components of fitness and so produce positive genetic correlations even when there are constraints on total fitness.
Balancing selection (e.g., due to heterozygote advantage) is more likely to lead to negative correlations, simply because the additive genetic variance for total fitness must be 0 if the only force acting is selection (pp. 462 and 548). However, given our ignorance of the causes of fitness variation, we cannot expect negative correlations in fitness components within populations.
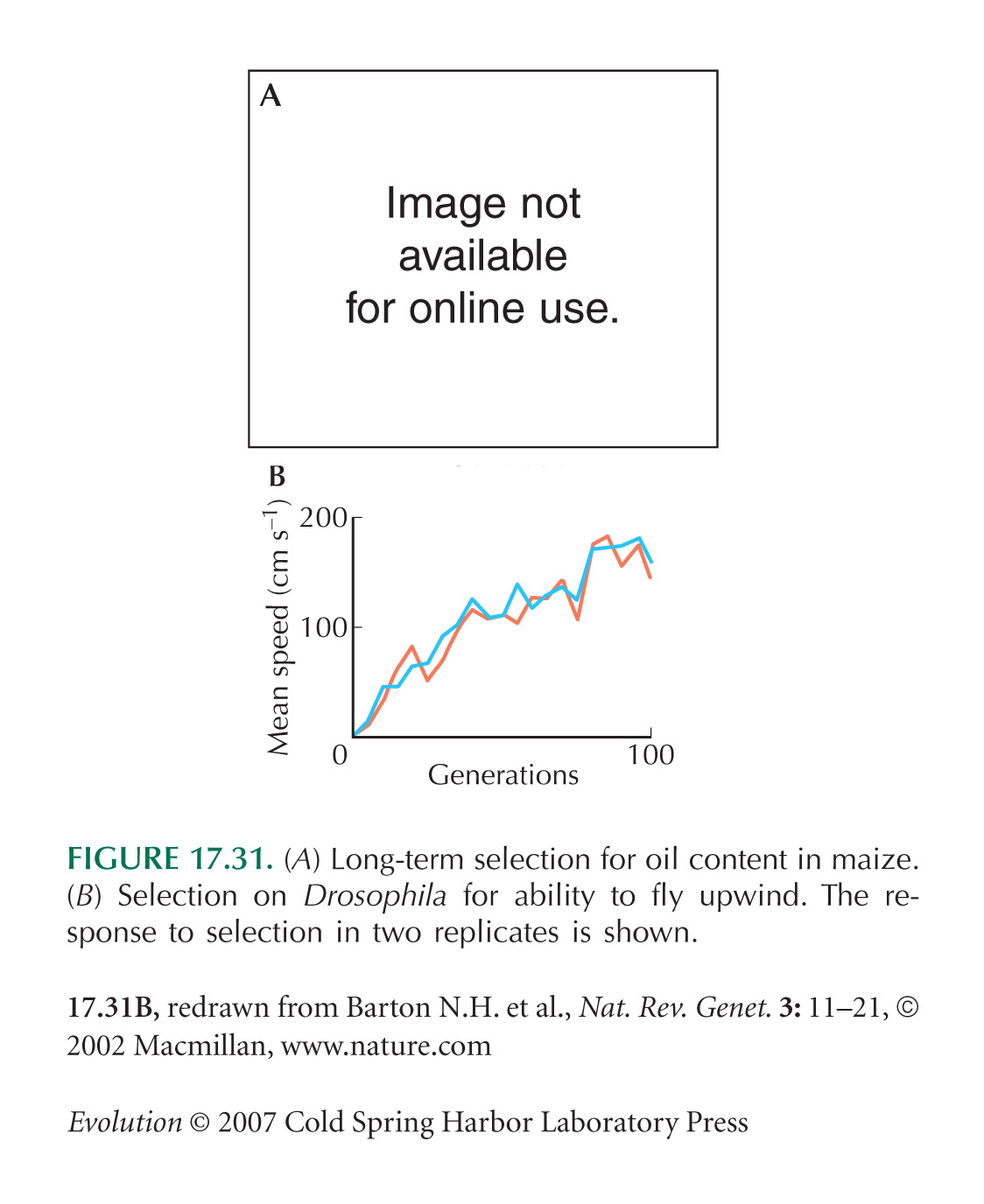
Artificial selection provides a more powerful way of exploring the constraints that shape organisms, because it can push the phenotype far beyond the range seen within any one population. This approach has been very successful in the study of aging, and in most cases has revealed the trade-offs that underlie evolutionary explanations of senescence (e.g., Box 20.2). However, evidence for trade-offs is not always found. For example, Weber achieved a remarkable increase in flight ability by selection on D. melanogaster for more than 100 generations (Fig. 17.31B). However, there was no evidence that these selected lines had lower fitness in other respects, such as competitive ability as larvae. Figure WN20.5 shows another example, the adaptation of E. coli to growth at different temperatures. After 2000 generations of experimental evolution, each population performed best at the temperature at which it had been selected, but there was little evidence for trade-offs. Fitness at different temperatures had not decreased, and the range of temperatures over which the bacteria could survive had not changed. Over the longer term, however, there is some evidence for trade-offs. After 20,000 generations at 37°C, E. coli populations did have lower growth rates at low and high temperatures (Cooper et al. 2001). Many experiments have been made on the evolution of host specialization, especially in insects. Here again, evidence for trade-offs is weak. For example, populations selected to feed well on one species of plant do not necessarily do worse on other plant species (see p. 653). Overall, the most important observation is that artificial selection can achieve remarkable changes without disrupting the functioning of the whole organism and without being much constrained by genetic correlations.
|



{kind=link}
{kind=link}
{kind=link}