Transposable Elements Can Spread Rapidly
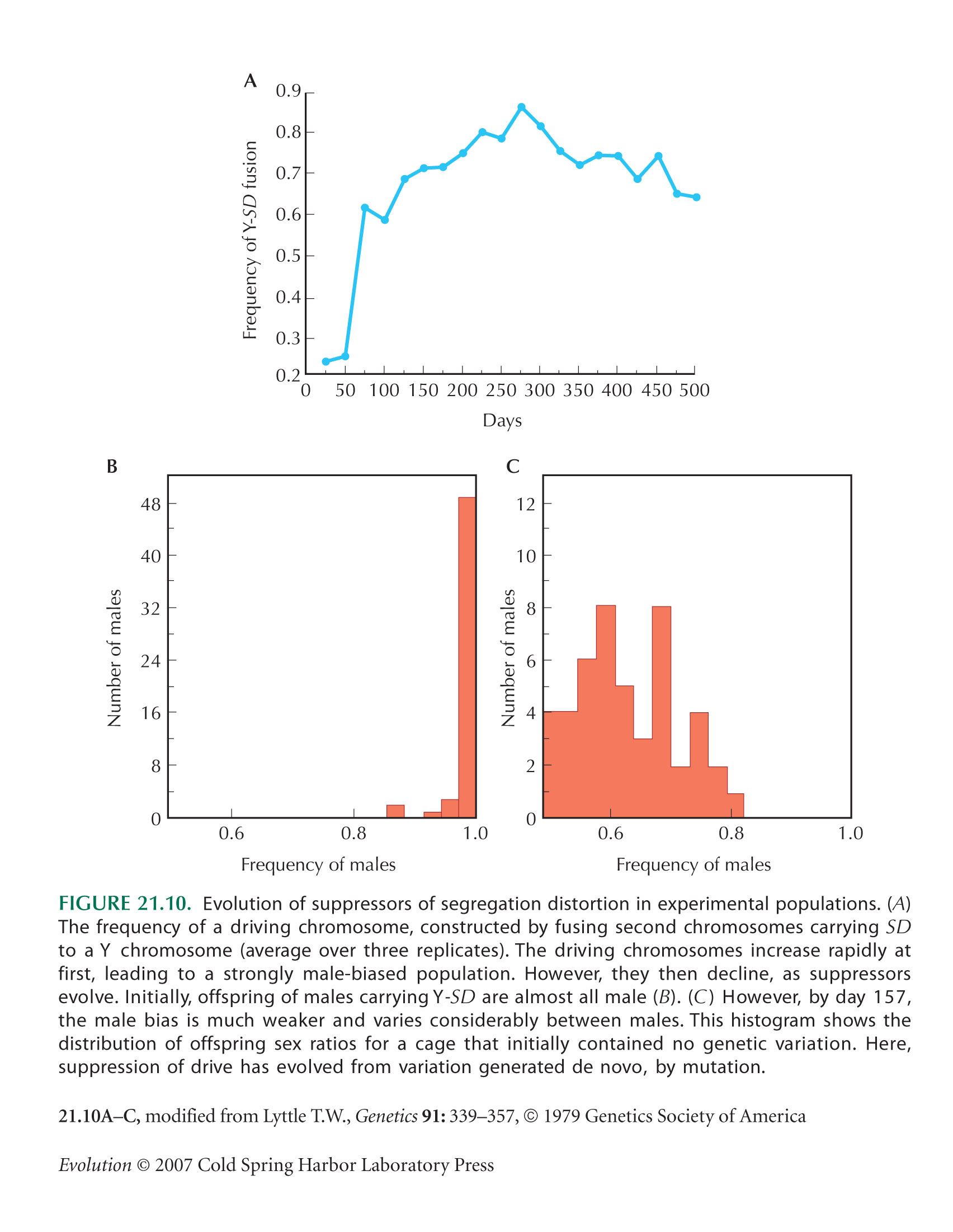
Genes with a transmission advantage can spread, even if they greatly reduce individual fitness. Indeed, the population may go extinct if it cannot maintain sufficient fitness despite these deleterious effects (at least two offspring must survive from each sexual parent). Driving sex chromosomes can cause extinction of one of the sexes (as, for example, in Lyttle’s experiments; Fig. 21.10). Transposable elements can spread to arbitrarily high copy number if the selection opposing each new copy is weaker than the rate of transposition.
The fixation or indefinite spread of selfish elements can be opposed only if the strength of selection against them increases with their abundance. The principle is the same as for any balanced polymorphism (see p. 506)—a stable equilibrium can be maintained only if there is some kind of frequency dependence. This may arise simply because the deleterious effect of each extra element becomes greater as they become more common. For example, a single B chromosome in the grasshopper Myrmeleotettix maculatus causes a barely detectable slowing of development, but two B chromosomes cause almost complete sterility (Hewitt and East 1978). In house mice, t/+ heterozygotes appear normal, but t/t homozygotes die early in embryogenesis or, if they survive, are sterile. In plants, male-sterile individuals lose fitness as they become common, because pollen then becomes scarce and they cannot self-fertilize.
Transposable elements can spread rapidly. One of the most remarkable examples is the P element, a DNA-based transposon found in Drosophila (pp. 342–343; Engels 1996, 1997). This is found in all natural populations of D. melanogaster, including the remote mountains of Central Asia. Yet, it is absent from laboratory stocks that were collected before the 1930s. The P element is spread widely across other Drosophila species (Fig. WN21.2), and the melanogaster element is almost identical to that in Drosophila willistoni (1 difference in 2907 base pairs), even though these species diverged about 60 Mya. This pattern is best explained if P elements spread throughout D. melanogaster very recently, after transfer from D. willistoni or a close relative. The willistoni group is endemic to Central and South America, and so spread must have begun sometime after European colonists brought D. melanogaster to the New World in the early 1800s.
Laboratory stocks that lack the P element are referred to as M strains. When P-bearing males are crossed to M females, offspring show hybrid dysgenesis: temperature-dependent sterility, high mutation rates, frequent chromosome rearrangement, and recombination in males. These phenomena are caused by mobilization of the P element in the germ line. (Artificially manipulated P elements that mobilize in somatic tissues cause high mortality. Presumably, restriction of transposition to the germ line, which is caused by an intron sequence within the transposase gene, is an adaptation by the P element to allow the host to survive, so that the element can be transmitted.)
The rapid spread of P elements into a population that has not been selected to resist them is atypical; usually, rates of transposition are low. For example, comparison of Drosophila lines that have been allowed to accumulate mutations gives an estimate of approximately 10–4 movements per transposon per generation (Maside et al. 2000). However, even this low rate would cause the number of copies to double over only approximately 7000 generations, indicating that it must be opposed by some kind of selection.
The suppression of transposition within populations carrying the P element is caused by accumulation of defective P elements, in which part or all of the transposase gene is deleted. These bind transposase but do not produce it themselves, and so reduce rates of transposition. Such elements are similar to the defective interfering viruses, discussed above. They can be looked at in several ways: (i) as parasites of the intact P elements, (ii) as an unselected by-product of transposition, or (iii) as suppressors of transposition that have spread because they reduce hybrid dysgenesis in their hosts. Many elements have unique defects, suggesting that they are by-products of failed transpositions. However, some (e.g., the KP elements) have spread to high frequency, which implies that they have been positively selected.
|



{kind=link}