Boundary Conditions for the Diffusion Equation
We can understand what happens at extreme allele frequencies (i.e., p = 0, 1) by thinking of the flux of probability (i.e., the net rate at which populations change their allele frequency). The diffusion equation (Eq. 36) for the allele frequency, p, can be written as
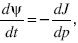
where
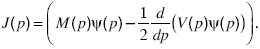
The quantity J(p) is called the probability flux. Imagine very many populations that follow the distribution of allele frequencies ψ(p). In a small time δt, some will increase their allele frequency from below a threshold, p, to above it, whereas others will decrease. The net proportion of populations that evolve from below p to above p is just Jδt. At equilibrium, this flux is zero; solving J = 0 gives the equilibrium distribution.
Suppose that a fraction A0 of populations have very low allele frequency, with p < p0 << 1, and a fraction A1 are almost fixed at very high allele frequency, with q < q1 << 1. Then, the rate of increase of A0 must equal the rate at which populations fall below p0 ~ 0:
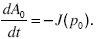
Similarly, the rate of increase of A1 must equal the rate at which populations rise above the threshold p1, which is close to 1:
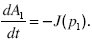
To understand these boundary conditions, think of a population evolving under mutation, selection, and random drift. The expected rate of change of allele frequency is M(p) = spq + µq – νp, where the rate of mutation from Q to P is µ and from P to Q is ν. The variance of allele frequency fluctuations is V(p) = pq/2Ne. Near the boundaries, selection is negligible relative to mutation and drift: For example, near p = 0, M ~ µ, and V ~ p/2Ne. Thus,
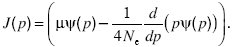
This has the solution ψ = C0 p4Neµ – 1. Thus, if the numbers of mutations entering the population are high enough (4Neµ > 1), there is a negligible chance of being at very low frequency. However, if 4Neµ < 1, the distribution becomes very large at the boundaries, implying that there is a high probability that one of the alleles will be lost. At equilibrium, J = 0, and so A0 stays constant. However, if the distribution is out of equilibrium, we can work out how the fractions fixed near 0 and 1 change through time.
|


