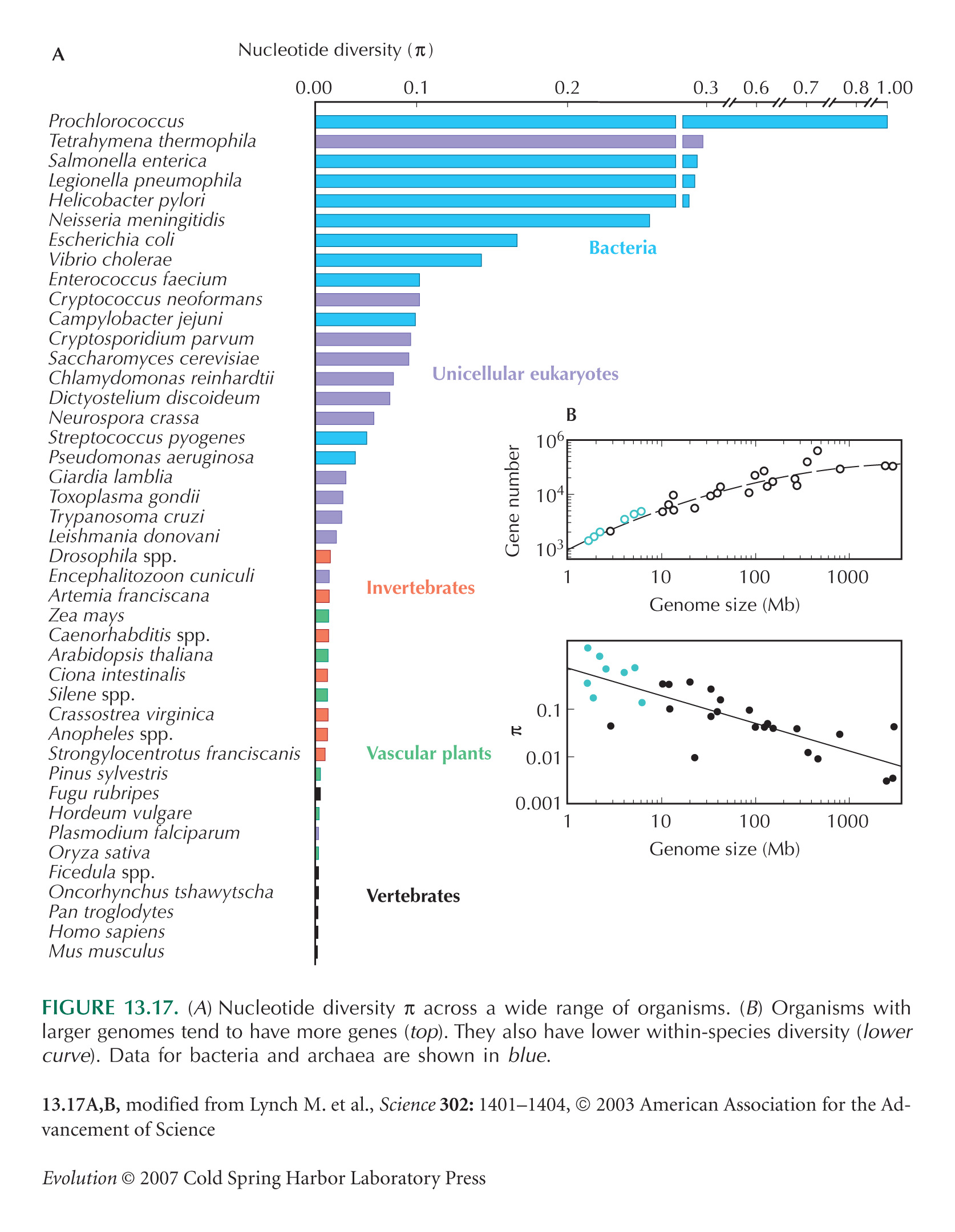
NOTE 18C: The nucleotide diversity is low in multicellular organisms (Fig. 13.17), and so this assumption can usually be made.
NOTE 18D: The frequency of preferred codons does not depend as strongly on population size as is implied by this calculation: Organisms with large Ne do not have preferred codon frequencies close to 1. See p. 544.
NOTE 18E: This prediction was derived by Clayton and Robertson (1955).
NOTE 18F: This seems an implausibly high value, given that eukaryotes typically have roughly 10,000 genes in total. This puzzle is discussed on pp. 409–411.
NOTE 18G: Selection is stronger than drift (Nes = 100 × 0.05 = 5) and so we can safely use the approximation P = 2s (see p. 493).
NOTE 18H: There is a subtlety here, when effective population size Ne differs from actual size N. The actual number of mutations generated at n loci is 2nNμ, but the probability that each one is fixed is 2s(Ne/N). Hence, the number of mutations that fix is 4snNeμ. Thus, we can use the effective poulation size throughout the calculation: The actual number cancels.
NOTE 18I: Hill (1982) gives the calculation of v), although with different notation. The agreeement between the two different approaches (one based on overall breeding value and the other based on fixation of individual alleles) depends on the assumption that alleles are equally likely to be positive as to be negative. Robertson (1960) gives a calculation that is based on fixation probabilities in a similar way, but assumes small Nes; this leads to the prediction 2NeβVm as in ii).
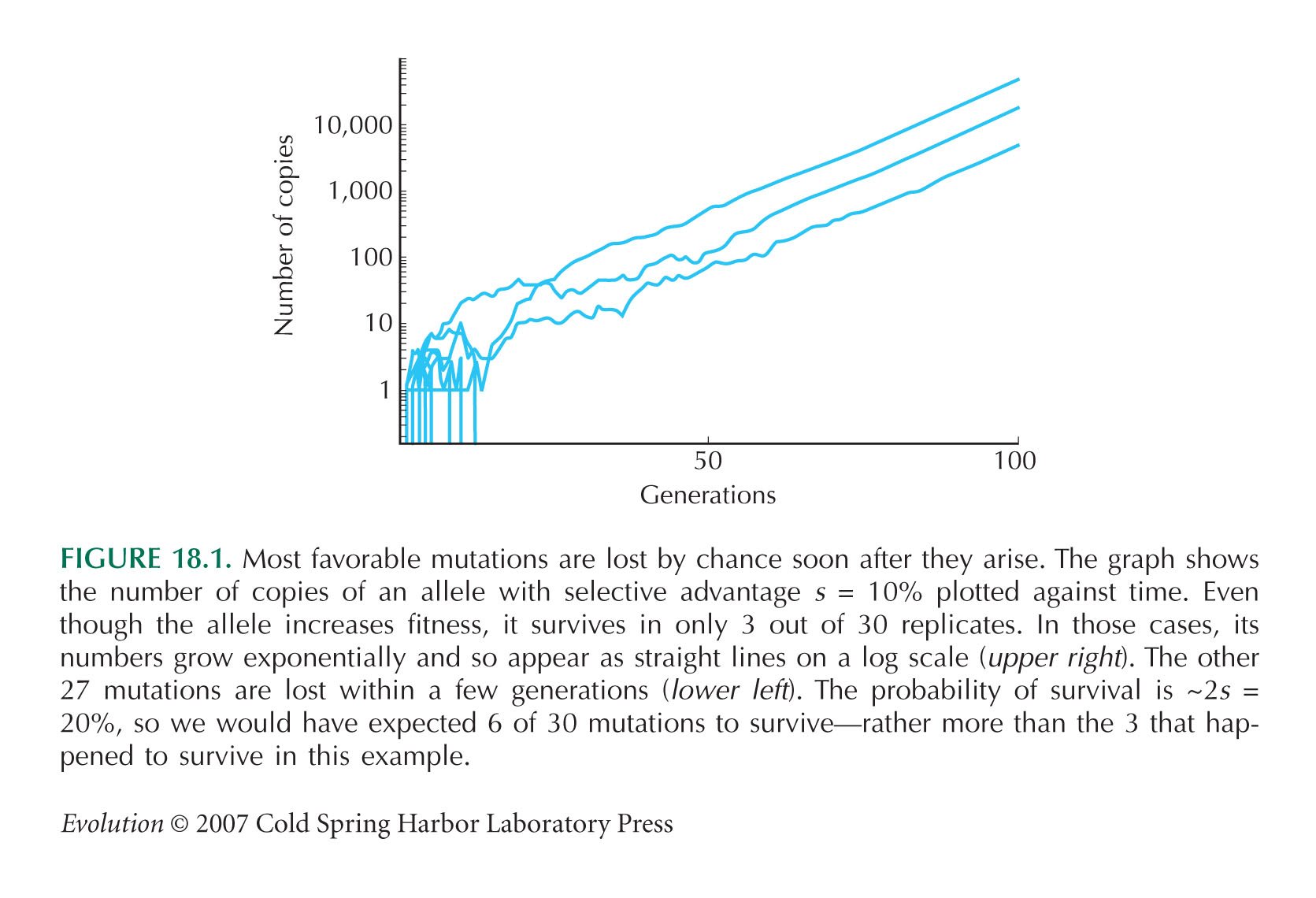
NOTE 18J: The deterministic expectation just described is actually an average over cases where the allele is fixed and where it is lost. The mutation has only a chance of 2s = 0.04 of being fixed, and so the expected frequency, given that it will be fixed in the population, is actually p0(1/2s)exp(st). (See Fig. 18.1.) This effect can be ignored here.
NOTE 18K: Conditional on fixation, the expected number of copies is (1/2s)exp(st).
NOTE 18L: The equilibria of this model were analyzed by Karlin and McGregor (1972).
NOTE 18M: When both alleles invade, we say that there is a protected polymorphism.
NOTE 18N: Although this question was set in terms of a haploid organism with two alleles at a single locus, it could equally well apply to two alleles in a diploid, with complete dominance (giving two phenotypes), or two asexual genotypes, or two species. The existence of different ecological niches (here, patches A and B), with competition within each, can maintain polymorphism as well as allowing different species to coexist. See p. 624.
NOTE 18O: We can see the answer to iii) using a simple mutation load argument: Every mutation that comes into the population must eventually be eliminated by a selective death. In every generation, 2Nμ mutations enter. If the selective deaths are of homozygotes, then each death removes two alleles, and we have 2 Nμ = 2NL, where L is the fraction that die because they carry the deleterious allele. See p. 552.
NOTE 18P: This is a version of the mutation load result (see p. 552). The loss of mean fitness due to genetic variance around the optimum is the average of z2/2Vs, which is VG/2Vs = U.
{kind=link}
{kind=link}