How Close to the Optimum Do We Expect Adaptations to Be?
Both Darwin and R.A. Fisher used the vertebrate eye as an example of a delicately optimized organ. There are good reasons to expect such a structure to be close to its optimum. First, parameters such as the shape and refractive index of the lens can be varied continuously by evolution, with no strong constraints. Second, the eye has a clearly defined function that is important for fitness. Similar arguments also apply to the example of dung fly mating behavior given in Box 20.1. In this case, the time spent copulating can vary freely, and mating is essential to male fitness.
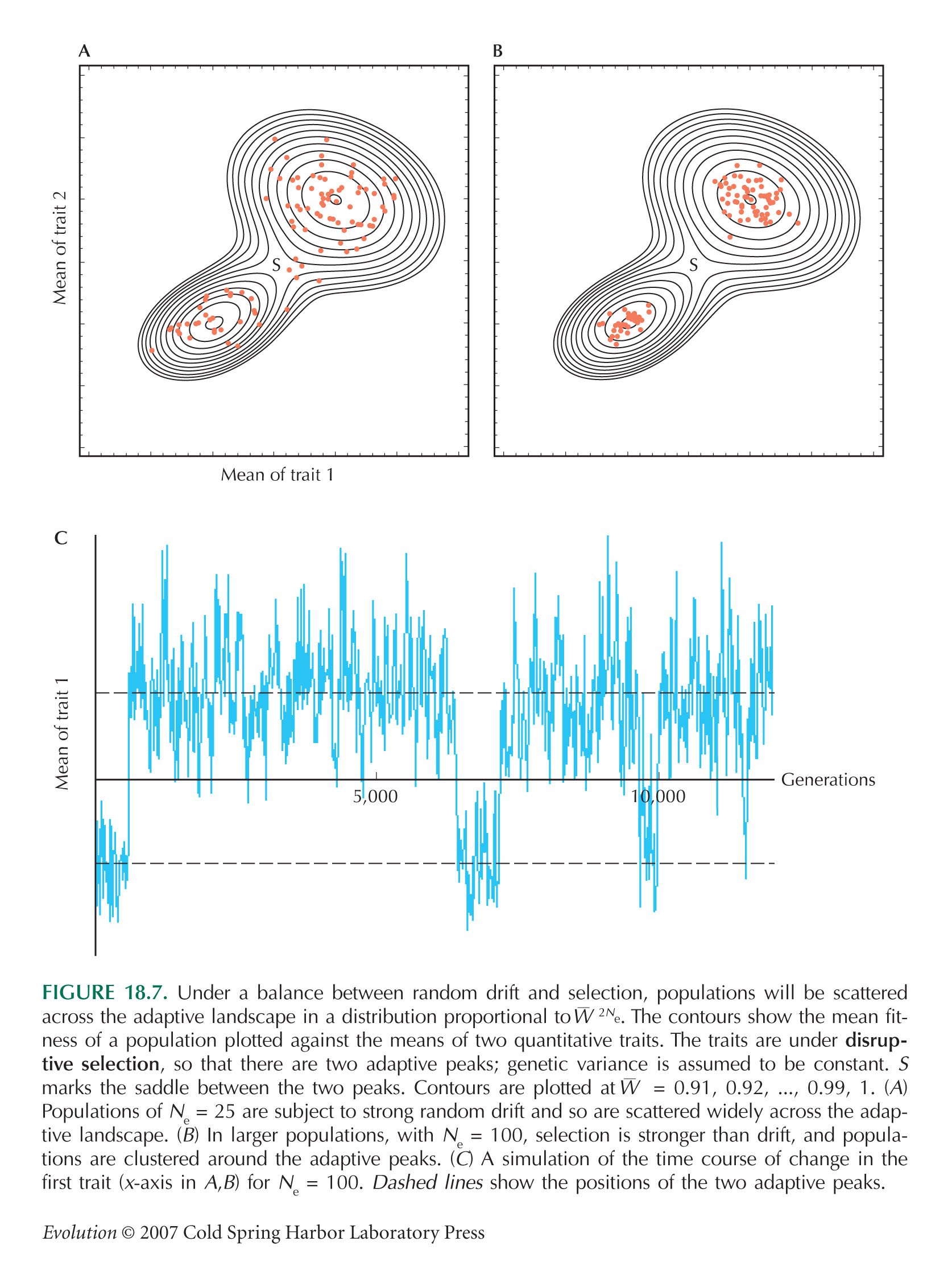
In contrast, we do not attempt to find an adaptive explanation for the detailed structure of genes. There are many qualitatively different ways of expressing genetic information, all of which can be adjusted to work well. To take a definite example, think of an RNA molecule that must maintain a paired stem–loop structure. Many different sequences can maintain essentially the same structure (Fig. WN20.2). To use Sewall Wright’s metaphor of an adaptive landscape, there may be many adaptive peaks (Fig. 18.7). Although an organism may be at a local optimum (e.g., a well-paired stem–loop structure), there may be many alternative, and more or less equivalent, optima, although which one is actually used will depend on the evolutionary history of the organism.
This contrast in the degree of optimization is not a contrast between molecular and organismal levels; there are many finely tuned molecular structures. We have seen that in highly expressed genes, and especially in microbes such as Escherichia coli and yeast, the triplet codon used to code for each amino acid tends to evolve to maximize efficiency and accuracy of translation (pp. 543–544). This molecular adaptation has evolved, even though selection on each codon must be weak.
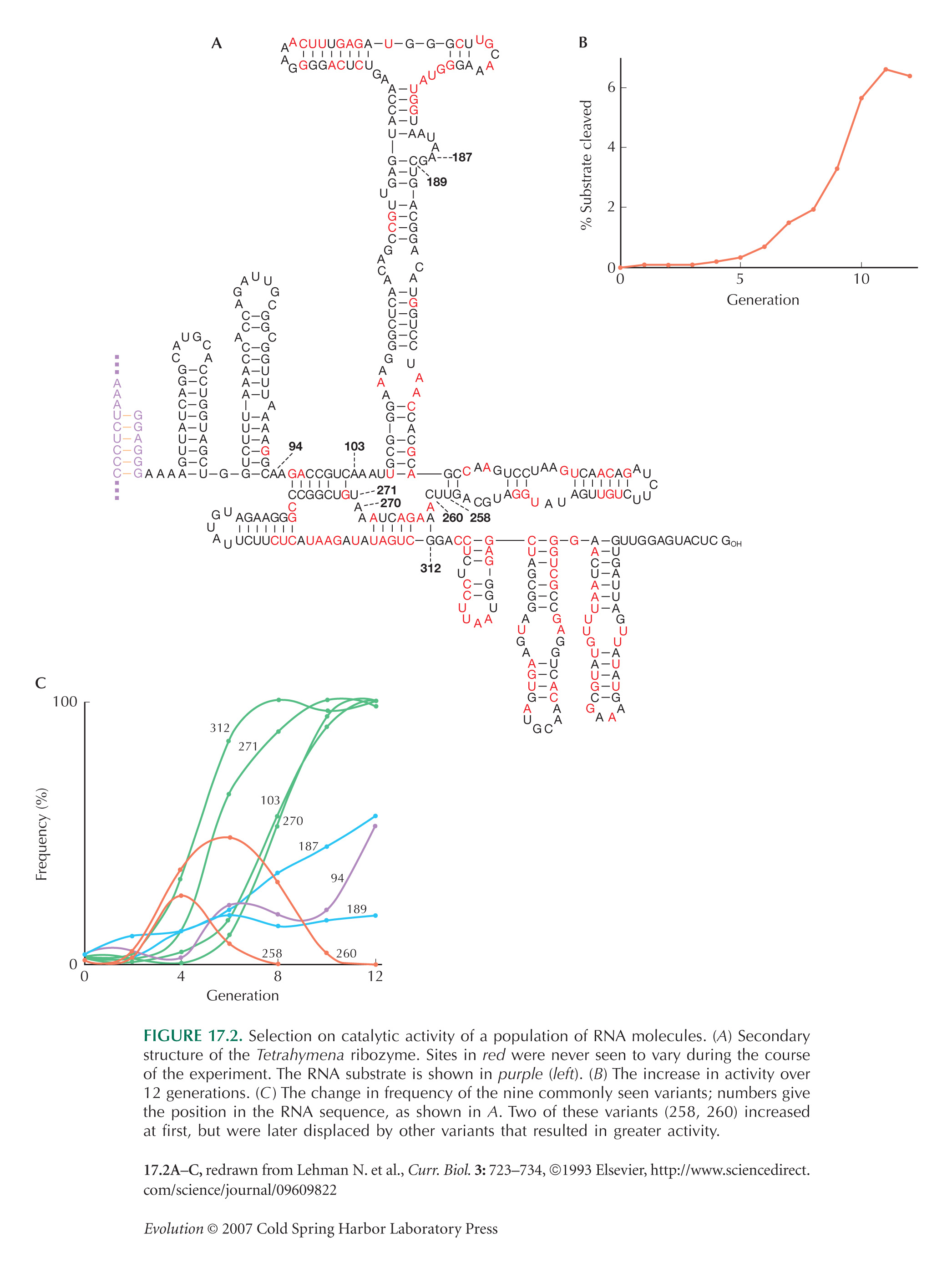
Artificial selection of RNA molecules gives us an exceptionally detailed understanding of molecular adaptation (recall Fig. 17.2). Here, both the fitness measure and the space of possibilities are known. Fitness is determined by the experimenter as the ability to catalyze a particular reaction or bind to a particular substrate, whereas the possibilities are the set of possible RNA sequences. It is impossible to explore all these possibilities. (Even a short molecule 40 bases long could have 440 [which is ~1024] possible sequences. This is the number of molecules in about 660 grams NOTE WN20A of single-stranded RNA.) However, as long as each local optimum can be reached from a wide range of similar sequences, the full range of possibilities can be explored in an in vitro selection experiment.
The hammerhead ribozyme is the smallest naturally occurring ribozyme, at about 40 bases long, and is found in plants, newts, schistosomes, and cave crickets. Its sporadic taxonomic distribution suggests that it has arisen several times by convergent evolution. This is supported by an elegant in vitro selection experiment by Salehi-Ashtiani and Szostak (2001; see also Doudna and Cech 2002) (Fig. WN20.3). They selected a random pool of sequences for the ability to self-cleave. After 12 rounds of selection, the pool was dominated by molecules that had similar sequences at the catalytic site (Fig. WN20.3C, magenta) and that had the complementary base pairing needed to form the correct secondary structure (Fig. WN20.3C, blue/yellow/red). Moreover, the activity of these newly evolved sequences was similar to that of the natural ribozymes (left column of Fig. WN20.3C). This is a rare example where the space of possibilities is small enough that selection can approach a single optimum from an entirely random starting point. In fact, many other types of ribozymes are found with a more efficient self-cleaving function, but all are considerably longer and thus are much less likely to be found in a short-term in vitro experiment.
Hermann Muller (1948) used dosage compensation in Drosophila (p. 682) as another example of how weak selection can make delicate adjustments to the genetic system. Most mutations are recessive and, indeed, most genes can be present in only one rather than two copies with no observable consequences. Hence, selection on the dosage of each gene cannot be strong. Nevertheless, Muller pointed out that Drosophila has evolved an elaborate mechanism of dosage compensation, such that genes on the X chromosome are usually expressed at twice the rate in males, which have only one copy of the X, relative to females, which have two copies. Dosage compensation can vary from gene to gene and so we see here an adaptation maintained by weak selection on the expression of individual genes. (Mammals have a different mechanism for achieving the same end: One X chromosome is turned off in females.)
There are many other examples where weakly selected traits are less closely optimized. The extreme case is the degeneration of vestigial organs, which was used by Darwin in On the Origin as strong evidence for evolution by natural selection. A nice quantitative example comes from West and Herre’s (1998) work on sex ratio in fig wasps (see p. 603). Selection favors an optimal ratio of males to females, in situations where a single female lays eggs on a fig. In species where single-foundress broods occur rarely, the sex ratio from single foundresses varies more than in species in which such broods are more common, so that selection is stronger.
There is a long-standing tension over the extent to which organisms can be expected to be “optimal.” This tension is reflected in the writings of two leading popularizers NOTE WN20B of evolutionary biology—Richard Dawkins, emphasizing the power of natural selection to produce adaptations, and Stephen Jay Gould, emphasizing that evolution is a historical process that depends on chance events (Fig. WN20.4). It is important to realize that the value of optimization arguments, of the kind explained here, does not depend on any assumption that organisms are in fact close to perfection. Rather, it is a way of understanding particular features of organisms: what selection favors, what constrains the possibilities, and how far selection dominates over other processes.
|



{kind=link}
{kind=link}