Island Radiations Suggest That Speciation Is Caused by Founder Effects
Ernst Mayr, Hampton L. Carson, Alan Templeton, and others have proposed various theories of founder effect speciation, which involve random drift driving severe population bottlenecks (see Carson and Templeton 1984; Barton and Charlesworth 1984; Barton, 1989, 1998). These theories were inspired by the remarkable diversification of species seen on many isolated islands. The best known example comes from the Hawaiian Drosophila, which includes at least 700 endemic species, compared to a total of approximately 1200 species in the rest of the world (see Carson and Kaneshiro [1976], and also the special issue of Trends in Ecology and Evolution, July 1987). The Hawaiian archipelago is more than 3500 km from the nearest continent or island group and was generated at a volcanic hot spot. Once formed, the islands were carried northwest by tectonic movement. Over time, they will erode and eventually disappear beneath the sea (Fig. WN22.8B). The phylogenetic relationships between the Hawaiian Drosophila species have been accurately reconstructed from the unique chromosomal inversions that they carry. Each species is endemic to one island and typically derives from an ancestor on an older island (Fig. WN22.8B). The history of these species must have involved multiple colonization events, most likely by a single fertilized female. The colonization events occurred on the archipelago as a whole, on each new island as it formed, and (within each island) on “kipukas,” which are patches of vegetation separated by lava flows. For reviews of Hawaiian radiations in several groups, including Drosophila, see Giddings et al. (1989) and Craddock (2000).
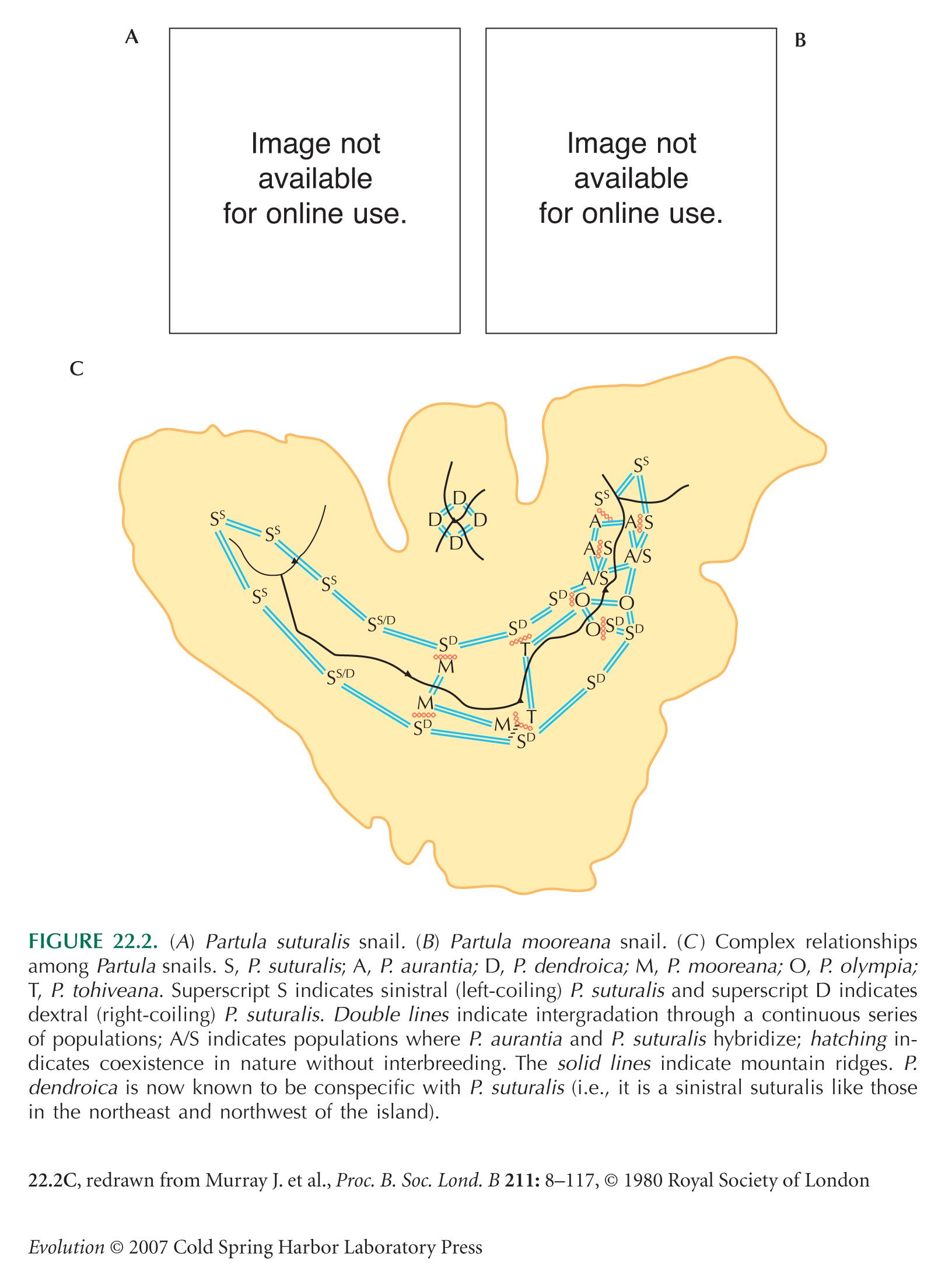
We have seen several other examples of island radiation, including Partula snails on Moorea (Fig. 22.2) and Geospiza finches on the Galápagos (Fig. 17.24). However, there is little evidence that such radiations have been driven by random drift during the founding of new colonies. Genetic variation within the Hawaiian Drosophila (measured by allozymes or by mtDNA) is similar to continental species. In Geospiza, there are multiple ancient MHC alleles that are shared across species. This implies that the Galápagos were colonized by a fairly large number of individuals, perhaps a stray flock (Vincek et al. 1997). Conversely, there are clear ecological factors favoring diversification. When a species colonizes an empty island, it has the opportunity to use ecological niches that would normally be exploited by other species. For example, the Hawaiian Drosophila occupy a much wider range of ecological niches than their mainland relatives. Most species live on the decaying parts of various endemic plants (leaves, fruit, and sap exudations). One group lives only on the flowers of the morning glory, whereas another is parasitic on spiders’ eggs.
Populations of flies in the laboratory have been subjected to repeated population bottlenecks to determine if this leads to reproductive isolation between replicate lines (Rice and Hostert 1993). Although weak assortative mating evolved in some experimental lines, overall there was no statistically significant evidence that bottlenecks strengthen prezygotic isolation. This contrasts with experiments where Drosophila populations have been selected to adapt either to new conditions or for changes in behavior or morphology. In these experiments, prezygotic and postzygotic isolations are often observed as a side effect of divergence. Overall, there is scant evidence for any role of random drift in speciation, either from laboratory results or from nature (Coyne and Orr [2004]).
|



{kind=link}
{kind=link}