Chapter 22 Notes
Species and Speciation
Introduction
Gould (1980) discusses the accuracy of folk taxonomies, referring to work by Berlin (1973). The example in the text is from Mayr (1963, p. 17).
The opening figure shows six of the bird species that coexist in the Sonoran Desert. The organisms that live together in one place usually cluster into distinct species.
Defining Species
The Continuity of Evolution Makes Species Hard to Define
Clear examples of ring species are hard to find. Examples are discussed by Irwin et al. (2001) and by Coyne and Orr (2004, Chapter 3).
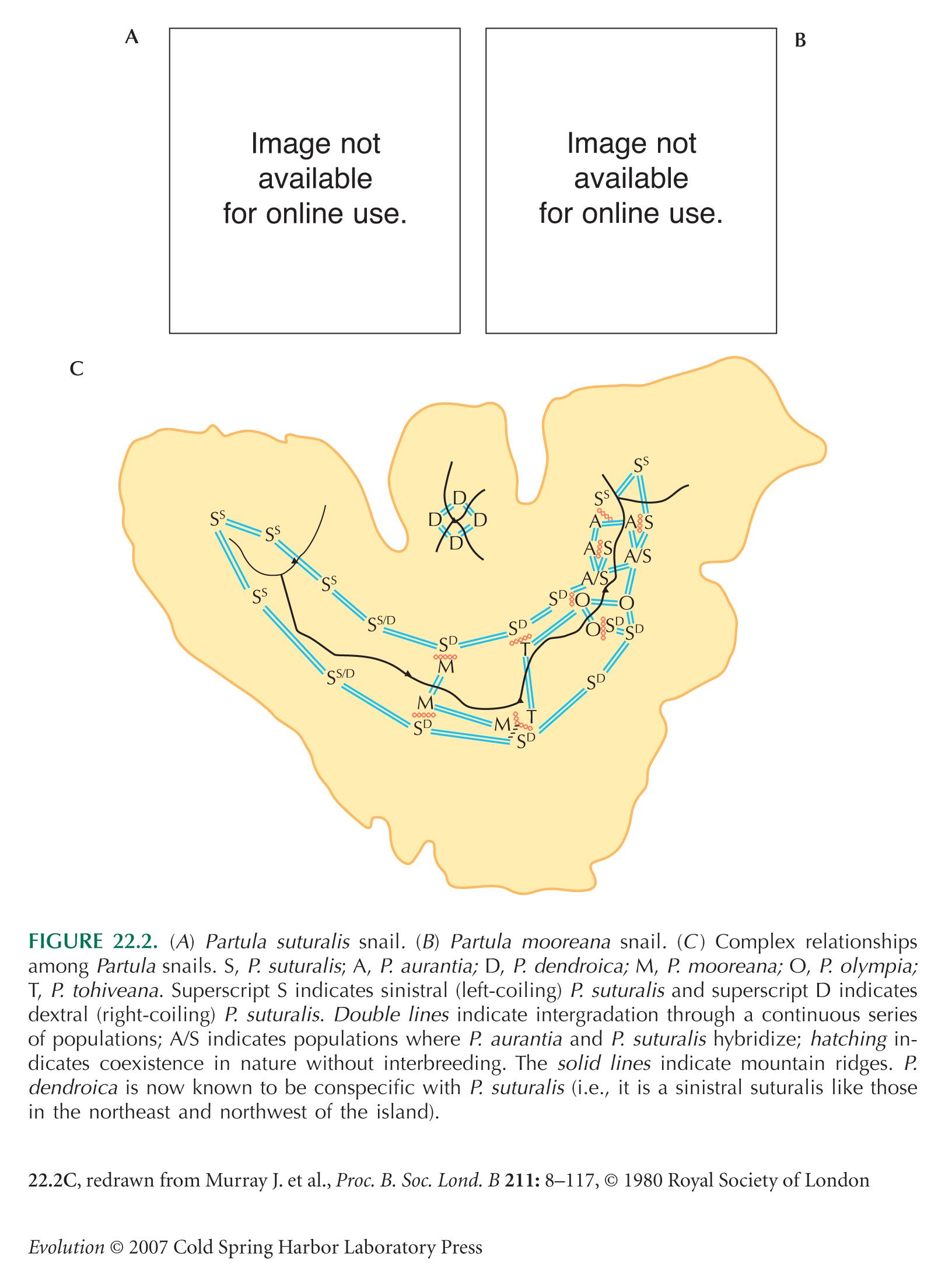
The complex relationships among Partula species (Fig. 22.2) are discussed by Murray and Clarke (1980). There was an error in Figure 22.2C in the first printing of the book: The M/T/S juncture at the bottom of the figure should show three hatched barriers, not two, because there is no evidence of hybridization between M and S there. In addition, P. dendroica is now known to be conspecific with P. suturalis (B. Clarke, pers. comm.). This error has been corrected in subsequent printings of the book and online.
There Are Several Ways of Defining Species
Desmond and Moore (1992, Chapter 23) give a detailed discussion of Darwin’s work on barnacles.
The microsporidian example of Figure 22.3 is from Katinka et al. (2001), discussed by Keeling (2001).
The cichlid example is detailed on pages 650–652, and in Web Notes. Verheyen et al. (2003) argue that the diversification in Lake Victoria occurred approximately 100,000 years ago, rather than after the present lake dried out 15,000 years ago, as had been thought previously. See also Seehausen et al. (2003).
Holder et al. (1999) show that there is substantial divergence in mtDNA between Indonesian and African coelacanths, suggesting that despite their morphological similarity, they may be separate species. Even so, the lack of diversity in present-day coelacanths and their similarity to ancestors that lived in the Devonian show a remarkably low rate of speciation.
The arbitrary figure of 97% sequence similarity for defining bacterial species is discussed by Gevers et al. (2005).
Barrett and Sneath (1994) define bacterial species by finding phenotypic clusters; see Cohan (2002) for further discussion of this approach.
Using Genealogies to Define Species Is Problematic
Hughes (1999) reviews evidence on sharing of MHC (major histocompatibility complex) alleles between species.
We explain the difficulties in defining species by their genealogy .
Harrison (1998) gives a clear discussion of the relationship between genealogical and biological species. Hudson and Coyne (2002) show that after two populations become reproductively isolated, it will still take a long time for the majority of their genes to share a genealogy that reflects this separation (i.e., to become genealogical species). Their paper is an interesting application of the coalescent process (see p. 422); see also Problem 22.2.
Biological Species Are Defined as Being Reproductively Isolated
The concept of the biological species is often associated with the Evolutionary Synthesis, and especially with Mayr and Dobzhansky. However, the idea goes back much further. Coyne and Orr (2004, Chapter 1) provide a good historical account and also include an appendix comparing different species concepts. Poulton (1908) gives an especially insightful discussion of what we now call the biological species concept, which refers back to Darwin’s views.
Whittemore and Schaal (1991) showed that chloroplast genealogies for white oaks do not match the morphology, but Howard et al. (1997) found that nuclear markers do, apart from regions where intermediates are found. Organelle genomes are often found to flow across species boundaries, as defined by conventional taxonomy or by nuclear genetic markers. This phenomenon is reviewed by Hudson and Coyne (2002), Rieseberg and Soltis (1991), and Fitzpatrick and Turelli (2006).
Figure 22.6B is from Howard et al. (1997) and Whittemore and Schaal (1991).
Species Can Coexist Only If They Use Different Ecological Resources
Hubbell (2001) has shown that a naive model of the random birth and death of ecologically equivalent species (analogous to the neutral theory of molecular evolution [p. 425]) fits observed distributions of species’ numbers surprisingly well. Ecologists have been skeptical of this argument, since species are plainly not precisely equivalent to each other. There is an interesting parallel here with the reception of the neutral theory, where the initial reaction was also that alternative alleles are unlikely to be precisely equivalent in their effects on fitness (pp. 59–60).
We discuss the fundamental question of why there are species (i.e., why organisms cluster into distinct populations) .
The biological species definition only applies to sexually reproducing organisms. , we discuss one way of defining species in asexual organisms, suggested by Cohan (2002). Gevers et al. (2005) review species definitions in bacteria, including Cohan’s.
Reproductive Isolation Can Arise in Many Ways
Mayr (1963, Chapter 5) provides a more detailed classification of causes of reproductive isolation than does Table 22.1.
An example of incompatibility with the maternal genotype comes from the cross between Drosophila simulans with D. melanogaster. When simulans males are crossed with melanogaster females, both sexes of the F1 hybrid survive. However, in the reciprocal cross, all females die as early embryos, because of an incompatibility between a maternally acting factor from Drosophila simulans mothers and a dominant X-linked factor from Drosophila melanogaster.
The firefly example of Figure 22.8B is reviewed by Lloyd (1966).
The cicada example is from Heliovaara et al. (1994).
Molecular Divergence Can Measure the Rate at Which Reproductive Isolation Evolves
Estimates of divergence times in the phylogeny of Figure 22.9 are from Tamura et al. (2004). They are calculated by averaging across many genes (62 for D. melanogaster vs. D. simulans) and correcting for the slower evolution of genes with strong codon usage bias (p. 543). However, no correction for polymorphism within species was made.
Klicka and Zink (1999) have argued that many bird species thought to have originated in the Pleistocene, in fact, diverged much earlier. Lovette (2005) reviews the subsequent debate and concludes that although some (especially boreal) species did originate in the Pleistocene, most diversification was earlier.
Reproductive Isolation Accumulates Slowly
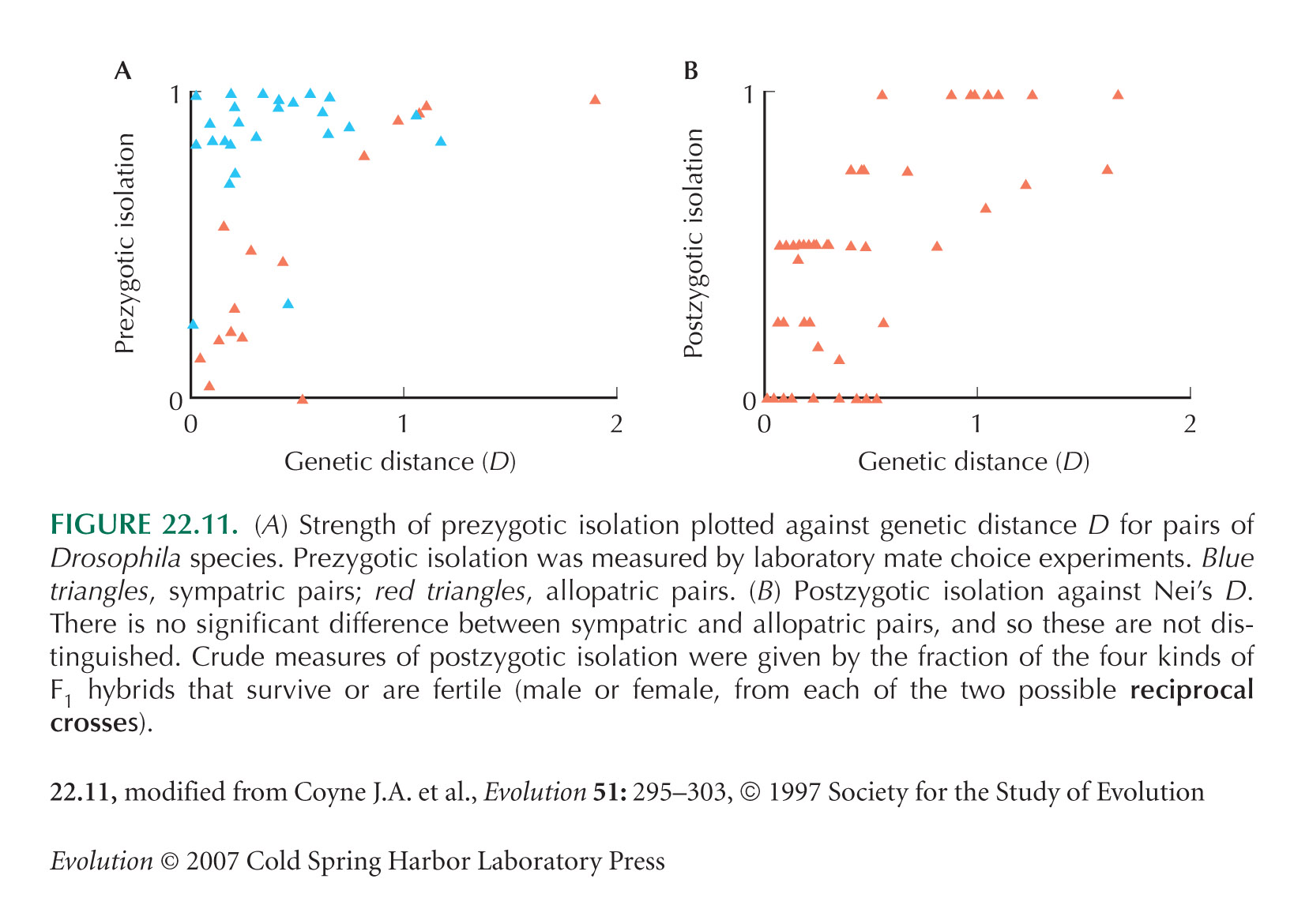
Surveys similar to Coyne and Orr (1997; Fig. 22.11) have been published for other groups. See Mendelson (2003), Presgraves (2002), Price and Bouvier (2002), and Russell (2003).
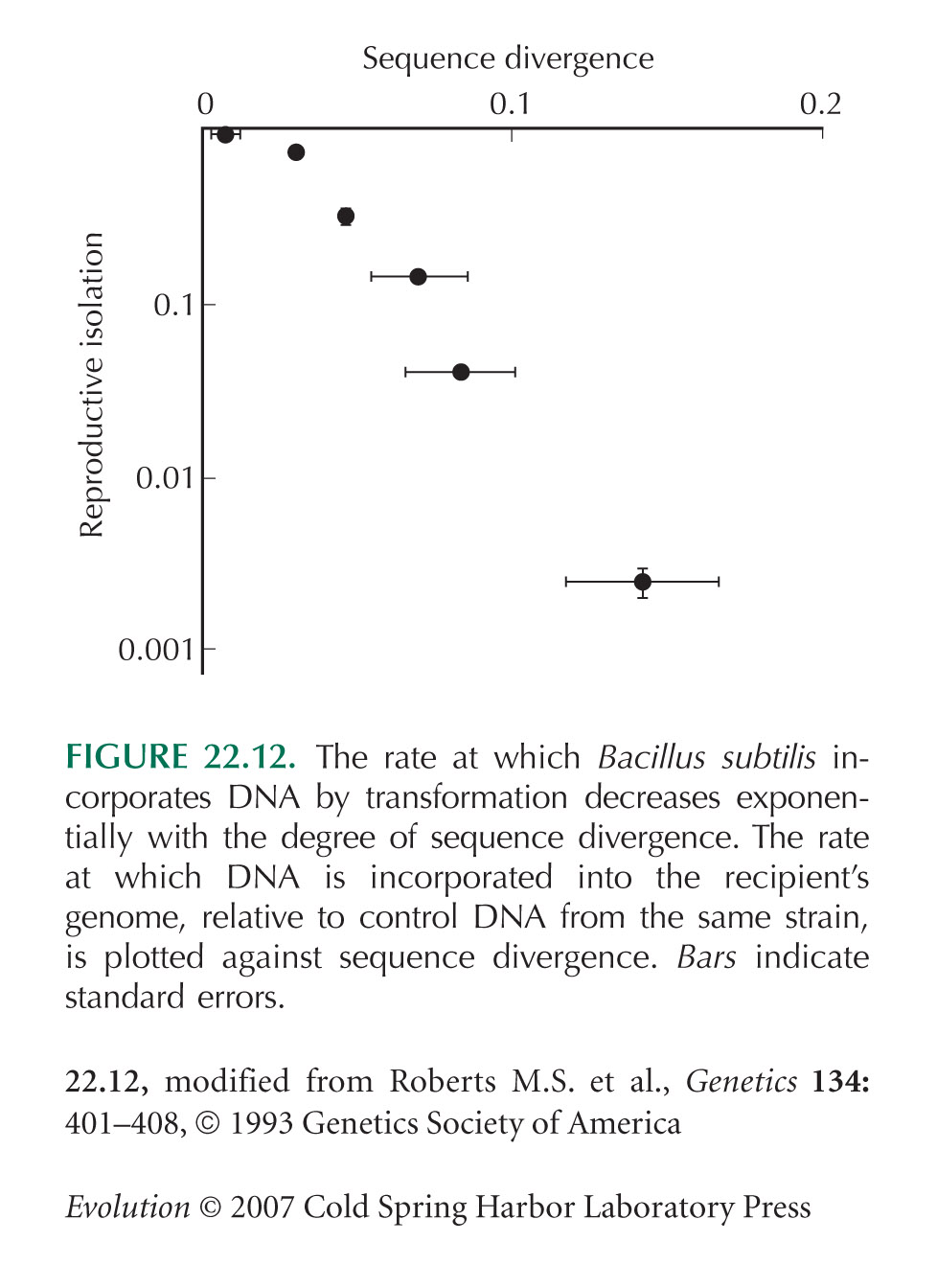
Figure 22.12 is from Roberts and Cohan (1993).
The Genetics of Speciation
Occasionally, Species May Be Separated by a Single Genetic Change
Occasionally, a new species can form from a hybrid population, without any change in ploidy. We discuss examples .
The Best-Understood Mechanism for the Origin of Species Is through Polyploidy
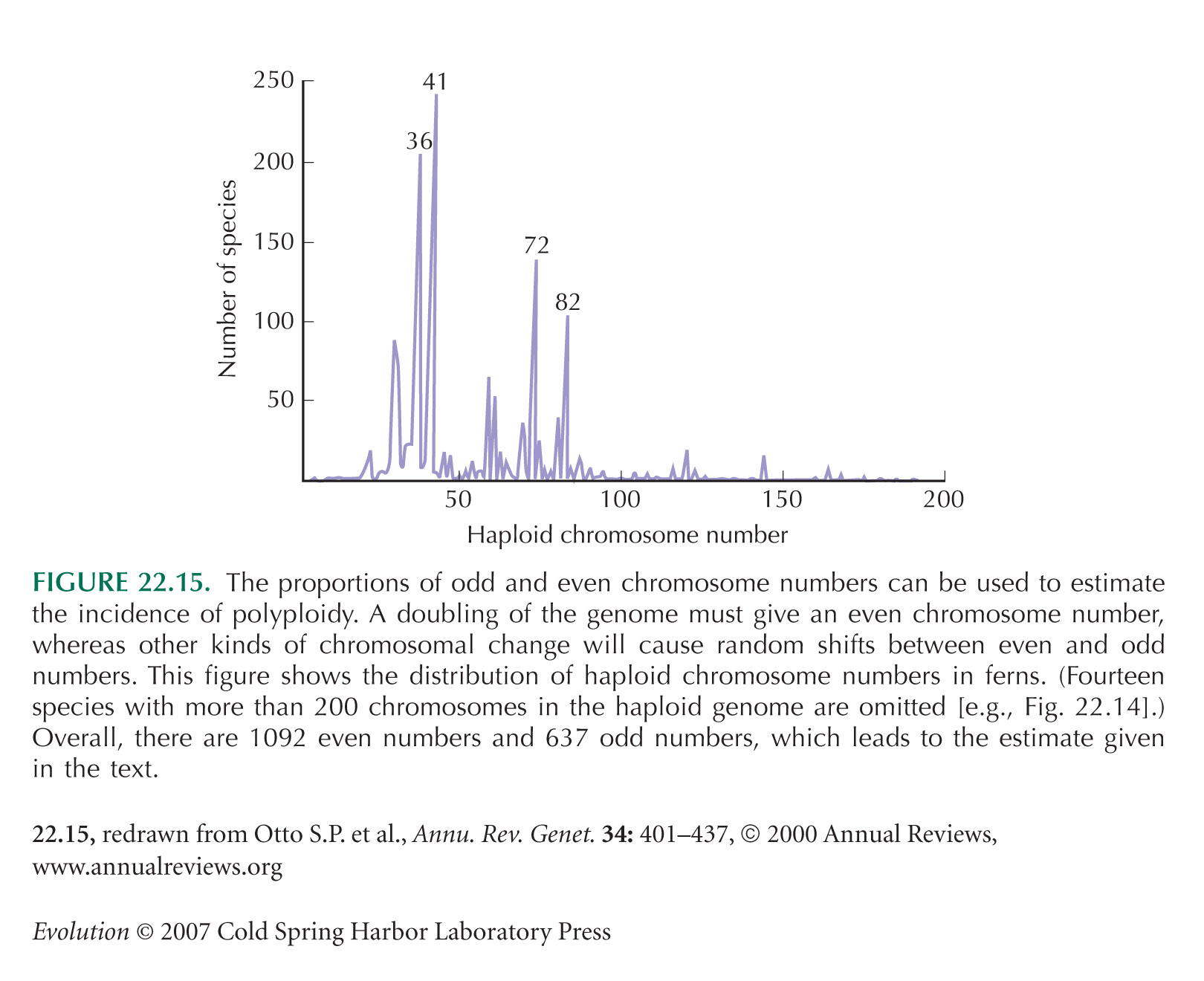
Otto and Whitton (2000) give an excellent recent review of polyploidy, including estimates of polyploid incidence made by comparing even and odd chromosome numbers (Fig 22.15).
Polyploids can form in several ways. Two sperm or pollen grains may fertilize the same egg, giving a triploid individual (polyspermy). A failure of mitotic cell division can produce tetraploid cells. In many organisms, an increase in ploidy is a normal part of somatic development (e.g., polytene chromosomes in flies are clearly visible because they consist of many copies, lying side by side; Fig. 13.31). If an extra doubling occurs in the germ cells, then diploid gametes may be produced. Finally, a failure of meiotic division can also lead to diploid gametes.
Muller (1925) suggested that an additional factor is that animals often have chromosomal sex determination, which may be disrupted by polyploidy. For example, an XXXY tetraploid individual would have three X chromosomes to every four chromosomes, rather than the usual 1:1 ratio in females or 1:2 in males. In organisms such as Drosophila, when the ratio of X to autosomes determines sex, this leads to intersexes. In addition, dosage compensation may be disrupted. Muller’s explanation is consistent with observations that most examples of sexual polyploid species in vertebrates are found in fish and amphibians, where sex is often determined by the environment (there are 10 examples among fish and 18 in amphibians). In contrast, there are virtually no examples in reptiles, birds, or mammals, where sex is usually determined chromosomally. However, in birds and mammals at least, polyploidy leads to general developmental problems, rather than specifically disrupting sex determination. In humans, for example, triploids and tetraploids rarely survive to birth, whereas sex chromosome abnormalities (XXY, XYY, etc.) cause relatively mild problems. See Orr (1990).
Figure 22.13 refers to Müntzing (1930).
For sources on highly polyploid plants (e.g., Fig 22.14), see Love et al. (1977) and Uhl (1978).
Evidence for ancient whole-genome duplications in the ancestry of vertebrates is discussed by Wolfe (2001), Seoighe (2003), and Lynch (2007).
Usually, Species Are Separated by Many Genetic Differences
Templeton (1977) gives an example of how the Wright–Castle method can be used to make a rough estimate of the numbers of genes responsible for species differences. See also Lande (1981) and Problem 14.7. As discussed on page 399, this method can do little more than show that “many” genes are involved. To accurately locate and characterize the actual QTL (quantitative trait locus) involved, we must follow the joint segregation of large numbers of genetic markers together with the trait.
For a detailed example of how variation in the gene Ubx is responsible for differences in bristle pattern between Drosophila species, see pp. 305–307 (Sucena and Stern 2000; Sucena et al. 2003).
Orr and Coyne (1992, Table 1) and Orr (2001, Table 1) review evidence on the genetic basis of species differences (but not including hybrid sterility and inviability).
For more on the Mimulus example, see Bradshaw et al. (1995, 1998) and Ramsey et al. (2003).
Classical Genetics Has Allowed Detailed Analysis of Reproductive Incompatibilities in Drosophila
The genetics of “speciation genes” are reviewed by Orr and Presgraves (2000), Coyne and Orr (2004, Chapter 8), Orr et al. (2004), and Orr (2005).
The only logical alternative to epistasis is that interactions are between the two copies of a gene at a single locus, so that heterozygotes suffer. Chromosome rearrangements apart (Box 12.2, Fig. 22.23), there is almost no evidence for this kind of incompatibility.
Orr and Irving (2001) dissect the genetic basis of incompatibility between Bogotá and USA populations of D. pseudoobscura. Orr and Irving (2005) show that F1 hybrid males are slightly fertile, and that the few offspring that they produce are all female. Thus, the incompatibility may well have been established through the fixation of an allele that causes segregation distortion, followed by evolution of suppressors (see pp. 587–593).
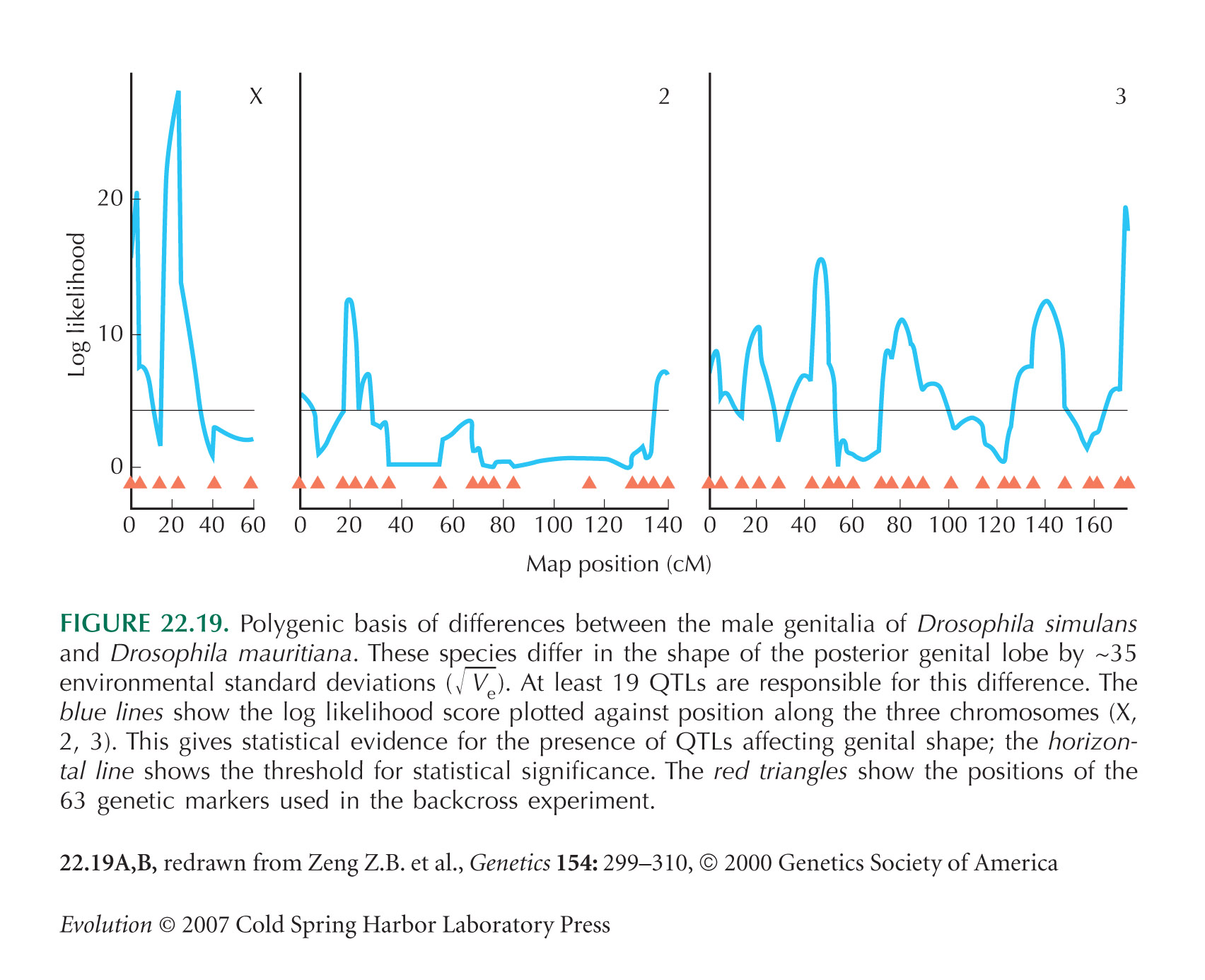
The genetic analysis of incompatibilities between D. mauritiana and D. simulans is from True et al. (1996). The genetic basis of differences in male genitalia between these species (Fig. 22.19) was determined by Zeng et al. (2000).
Hybrid rescue alleles in Drosophila are analyzed by Sawamura (1993a,b), Barbash et al. (2000), and Orr and Irving (2000).
Some of the Genes Responsible for Reproductive Isolation Have Been Identified
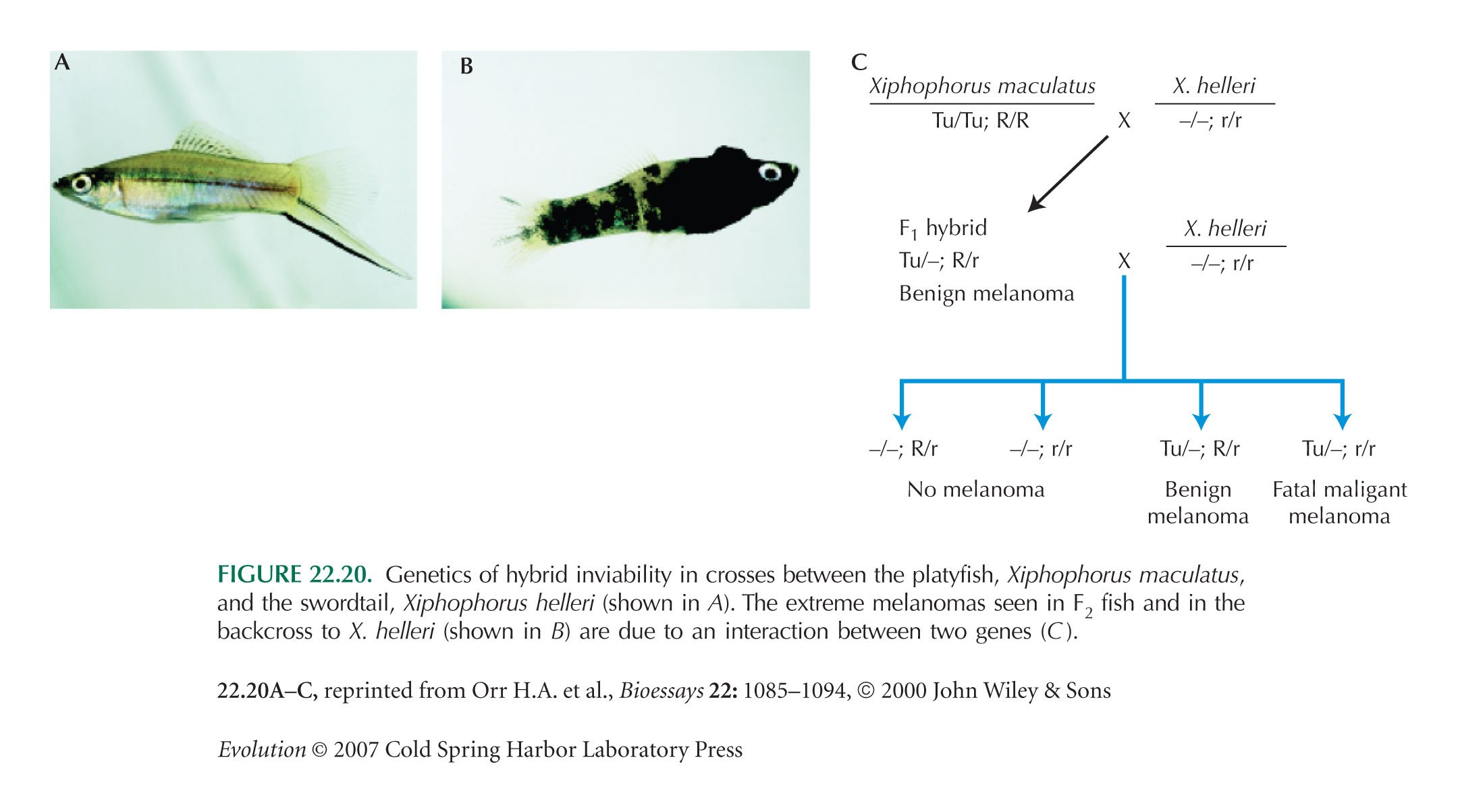
In Xiphophorus maculatus, Tu contains three tightly linked genes: Mdl (melanocyte determining locus), Xmrk-1, and Xmrk-2. The gene Xmrk-2 is a duplicate of Xmrk-1 and is nearly identical to it, but it is controlled by a different promoter. It is found on the X chromosome of X. maculatus but not on the Y chromosome or in X. helleri. The level of expression of Xmrk-2 transcripts correlates closely with the size of melanomas and is controlled by the second locus (R/r). This locus is thought to be CDKN2X, a cyclin-dependent kinase whose homologs in humans and rats act as tumor suppressors. Orr and Presgraves (2000) give a good account of the Xiphophorus example (Fig. 22.20).
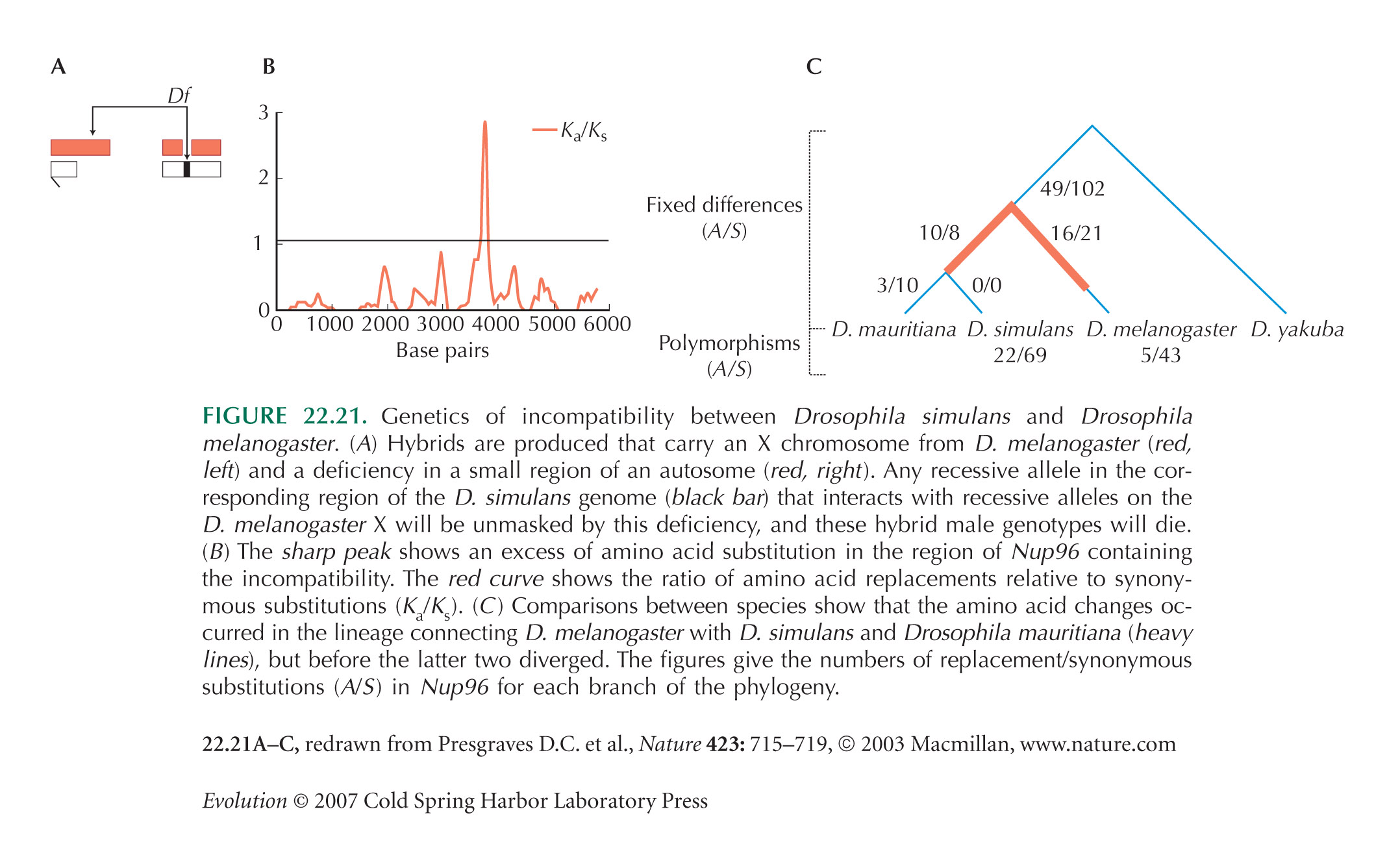
Presgraves (2003) describes the screen that detected incompatibilities between D. simulans and D. melanogaster. Presgraves et al. (2003) detail one of these, Nup96 (Fig. 22.21). This is surprising, because this gene codes for an essential component of the nuclear pore complex that is responsible for transporting mRNA out of the nucleus. Presgraves and Stephan (2007) show that the proteins with which the Nup96 product interacts also have undergone adaptive evolution, suggesting rapid coevolution between these genes.
Haldane’s Rule Tells Us about the Genetic Basis of Species Differences
Haldane (1922) first described Haldane’s rule. Muller (1940, 1942) outlined the dominance theory, but the explanation only became clear in the 1990s. For a review, see Orr (1997a).
The figure for D ~ 0.26 is the weighted average of classes 0.25, 0.50 in Table 2 of Coyne and Orr (1997).
Read and Nee (1991) have pointed out that the data in Table 22.2 are not compelling evidence that it is heterozygosity of the sex chromosomes that is responsible for the asymmetry in hybrid fitness between the sexes. Female sex chromosome heterozygosity evolved independently in birds and Lepidoptera, and so strictly speaking, there are only two independent comparisons here (Chapter 27). The pattern could, in principle, be due to some other feature of birds and Lepidoptera, which just happens to coincide with their different patterns of sex determination. However, Haldane’s rule is obeyed in several other groups such as lizards, salamanders, and stick insects. To date, seven independent associations between sex chromosome heterozygosity and hybrid sterility/inviability are known (see Orr 1997a). In addition, we will see that the likely explanation for Haldane’s rule directly implicates the sex chromosomes.
Haldane’s rule provides good evidence that most incompatibility alleles are recessive. Thus, in species having heterozygous sex chromosomes in males, the F1 males will be inviable or sterile if a dominant allele arises on the autosomes and interacts with any incompatibility alleles (recessive or dominant) on the X. In contrast, F1 females only suffer when two or more dominant incompatibility alleles interact. Because dominant alleles are much rarer than recessive, F1 female sterility or inviability takes much longer to evolve.
A simpler test of the dominance theory uses whole chromosomes. When D. simulans and D. melanogaster are crossed, F1 males die, but F1 females, which carry one X from each species, survive. Female hybrids can be constructed that carry two D. melanogaster X chromosomes or two D. simulans X chromosomes. These die at the same stage as the F1 males and, moreover, can be rescued by the same hybrid rescue allele as F1 males (see ). (This test was first carried out by Coyne [1985], who crossed D. simulans with D. sechellia and D. mauritiana. He found that although F1 males were sterile, hybrid females with two identical X chromosomes were fertile. At the time, this was thought to refute the dominance theory. Later, it was realized that because different genes affect male and female fertility, one does not necessarily expect females to be sterile in this test. Making an X that carries recessive alleles for male sterility homozygotes will only give sterile females if it also happens to carry recessive female sterility alleles.)
Haldane’s Rule Predicts the Large X Effect
Charlesworth et al. (1987) review evidence for the large X effect and discuss possible explanations. However, they focus on reasons why the actual rate of evolution might be faster on the X rather than on reasons why incompatibilities on the X are more likely to be expressed, as postulated by the dominance theory.
Turelli and Begun (1997) survey the effects of X-chromosome size in Drosophila.
True et al. (1996) found sterility factors more often on the X than on the autosomes, which suggests that incompatibility evolves faster on the X than on the autosomes (rather than simply being uncovered there, as under the dominance theory). However, True et al. add the caveat that the introgressions were longer on the X than on the autosomes, so this is not compelling evidence of a real difference.
Presgraves and Orr (1998) compare Haldane’s rule in Anopheles and Aedes mosquitoes (Table 22.3).
Payseur et al. (2004) argue that steep clines reflect strong selection for X-linked markers in the hybrid zone between Mus musculus musculus and M. m. domesticus (see ). However, Macholan et al. (2007) reanalyzed their data and did not find systematically narrower clines for X-linked markers. In a different transect, Vanlerberghe et al. (1986, 1988) do find a remarkably narrow cline for Y-linked markers, clearly indicating strong selection.
Masly and Presgraves (2007) introgress small segments of the D. mauritiana genome into D. sechellia and find evidence for both the dominance theory and for faster evolution of male than female sterility. They also find an excess density of male steriles on the X chromosome, which is responsible for the large X effect in this species pair.
Mechanisms of Speciation
Random Drift Can Knock Populations from One Adaptive Peak to Another
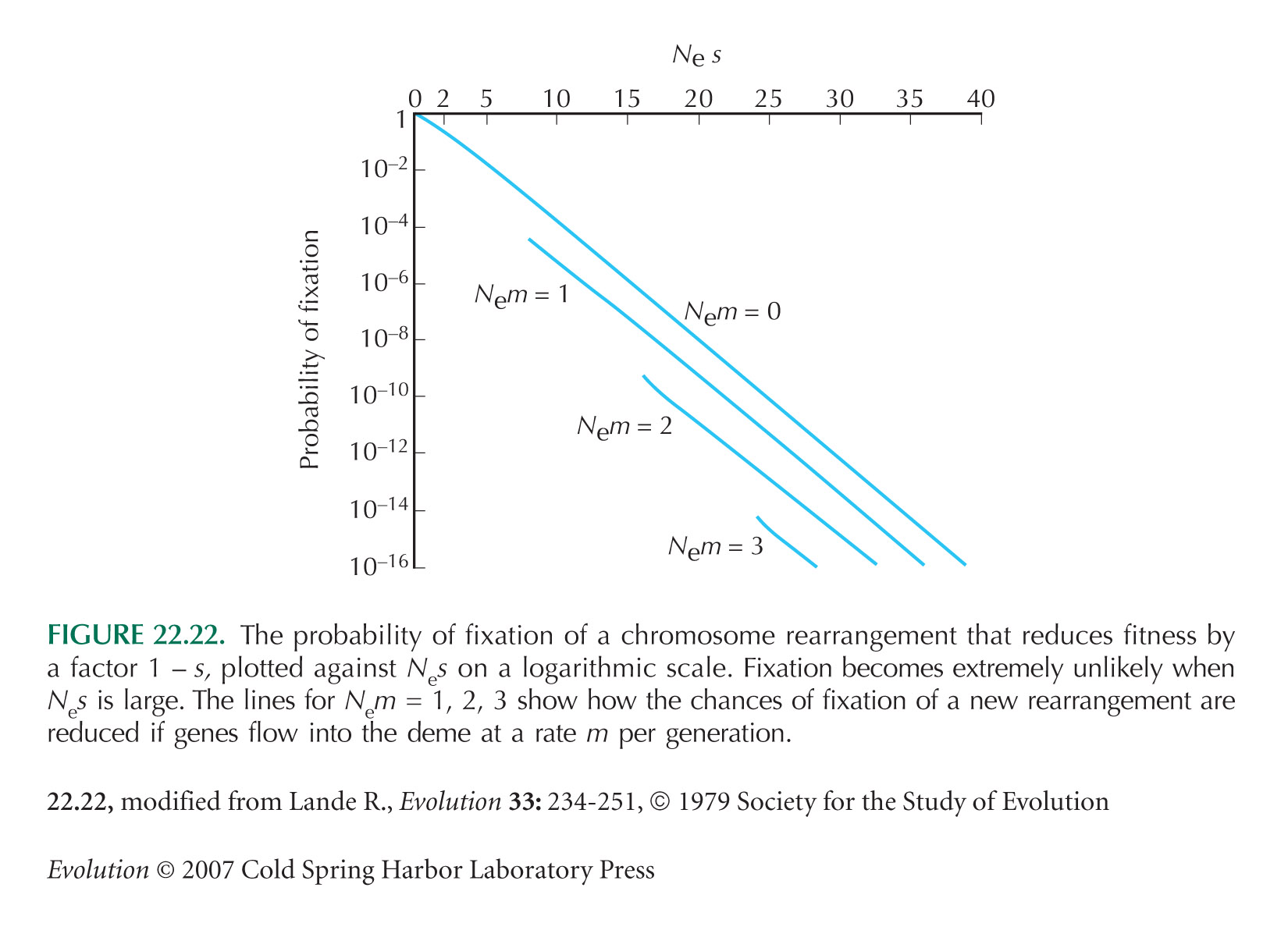
Wright (1941) first calculated probabilities of fixation of chromosome rearrangements. Lande (1979, 1984, 1985) extended Wright’s argument to show how rearrangements could spread through subdivided populations. Figure 22.22 is from Lande (1979).
The argument that drift is most likely to build up reproductive isolation through peak shifts of strength ~1/Ne was made by Walsh (1982).
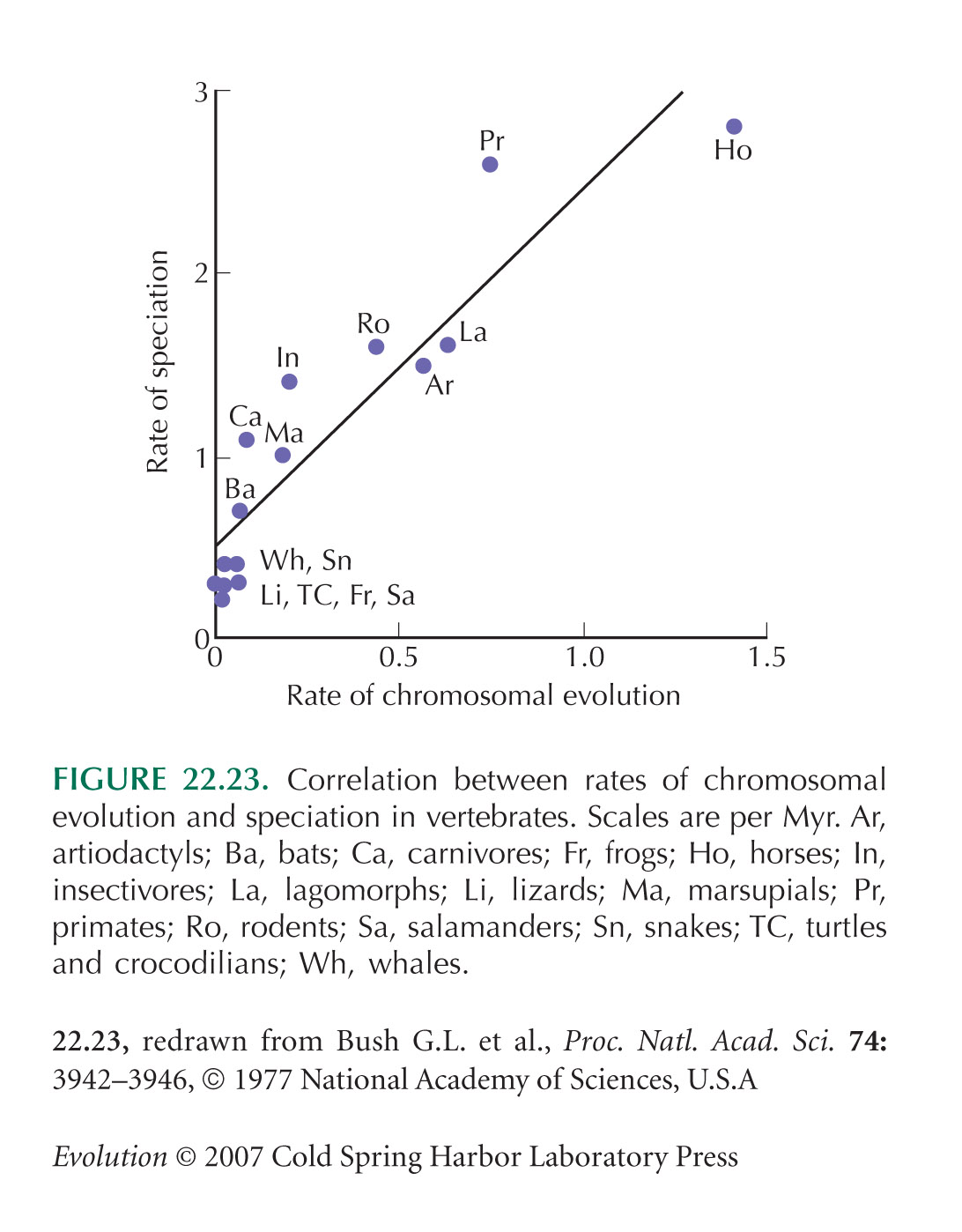
Evidence that rates of speciation are correlated with rates of chromosomal divergence (Fig. 22.23) is from Bush et al. (1977), Wilson et al. (1975), and Levin and Wilson (1976). These correlations between speciation rate and chromosomal divergence are quite crude. Speciation rate per lineage per Myr is calculated from numbers of species, assuming binary branching; only chromosome changes that alter chromosome numbers are included; and the significance of correlations is tested without taking into account phylogenetic correlations. However, these assumptions are unlikely to much affect the strong overall pattern.
The origin of large numbers of species on oceanic islands suggests that speciation is caused by founder effects, although a diversity of empty niches may also be important. We discuss this issue .
Reproductive Isolation Can Evolve without any Transient Reduction in Fitness
Gavrilets (2004) has argued that populations are most likely to diverge along ridges of high fitness, rather than by crossing deep valleys in opposition to selection. His book (2004) reviews these theoretical ideas and the evidence that supports them.
In the Dobzhansky–Muller Model, Speciation Is Not Opposed by Selection
This model was proposed independently by Dobzhansky (1936) and by Muller (1940, 1942), but had first been proposed by Bateson (1909). Orr (1997b) reviews the history of these ideas.
The shrew example (Fig. 22.26) is from Searle (1986). Amelioration of chromosomal incompatibilities is also seen in mice (M, musculus domesticus; see Searle 1993). Virdee and Hewitt (1992) and Shuker et al. (2005) have demonstrated a similar amelioration in a hybrid zone between two subspecies of the grasshopper Chorthippus parallelus (see Chapter 19 Web Notes). F1 males from crosses between these subspecies are sterile—an instance of Haldane’s rule. However, no such sterility is seen in nature, showing that intermediate genotypes with high fertility occupy hybrid populations. This shows that these incompatible subspecies could have diverged via a series of fertile intermediates, without opposition from selection.
Kondrashov et al. (2002) show that, frequently, amino acids that cause lethal disease in humans are fixed in other species. This provides another way to observe the kinds of incompatibilities proposed in the Dobzhansky–Muller model. We describe these results . Gao and Zhang (2003) discuss possible objections to the interpretation of Kondrashov et al., but find these objections implausible. Kulathinal et al. (2004) use a slightly different method to identify compensated pathogenic deviations (CPDs) using comparisons with Drosophila and find a similar prevalence. Kern and Kondrashov (2004) extend the survey to show that mitochondrial tRNA variants that are lethal in humans are often fixed in other mammals.
The Geography of Speciation
Speciation Does Not Require Geographic Barriers
Coyne and Orr (2004, Chapter 3) give a good historical account of the geography of speciation; see also Fitzpatrick and Turelli (2006).
The example of Figure 22.28B is from Pounds and Jackson (1981). Morphological divergence is based on 28 skeletal traits.
In the text, we argue that concordant clines indicate secondary contact. The alternative is that selection acts in the same way on many genes and traits and that the selection is so strong as to prevent exchange of neutral genetic markers. This seems implausible in most cases and can be refuted by observing the actual rate of gene flow.
The degree of overlap between the ranges of the most closely related species has been used to indicate whether they arose in allopatry, parapatry, or sympatry. Barraclough and Vogler (2000) survey range overlap plotted against degree of divergence. Fitzpatrick and Turelli (2006) survey mammal species and find more cases of range overlap than did Barraclough and Vogler, giving weaker evidence for allopatric divergence.
Navarro and Barton (2003) show how barriers to gene flow can promote divergence under the Dobzhansky–Muller model. This is explained in Figure WN22.10.
The argument that speciation does not require absolute geographic barriers but may instead occur in parapatry was made by (among others) Endler (1977). See also Coyne and Orr (2004, Chapter 3) and Gavrilets (2004, Chapter 11).
For opposing views on whether clines evolved in situ (primary contact) or instead are due to secondary contact, see Endler (1982) and Mayr and O’Hara (1986).
Genealogies and Chromosomal Rearrangements Give Evidence for Gene Flow during Speciation
Rieseberg and colleagues have investigated the genetic factors that keep Helianthus annuus and H. petiolaris separate. These two species meet in narrow hybrid zones, less than 50 meters wide, in Nebraska. Plants in hybrid populations that were morphologically similar to H. annuus are likely to have been produced by multiple generations of backcrossing to H. annuus. As expected, H. petiolaris markers were rare in such backcrossed hybrids. Crucially, H. petiolaris markers were only half as frequent in regions of genome that had different chromosome arrangements, compared with nonrearranged regions (13.1% vs. 7.1%). Moreover, the QTLs responsible for pollen sterility tend to lie in those same rearranged regions, in both wild and experimental hybrids (Fig. WN22.4). See Buerkle and Rieseberg (2001).
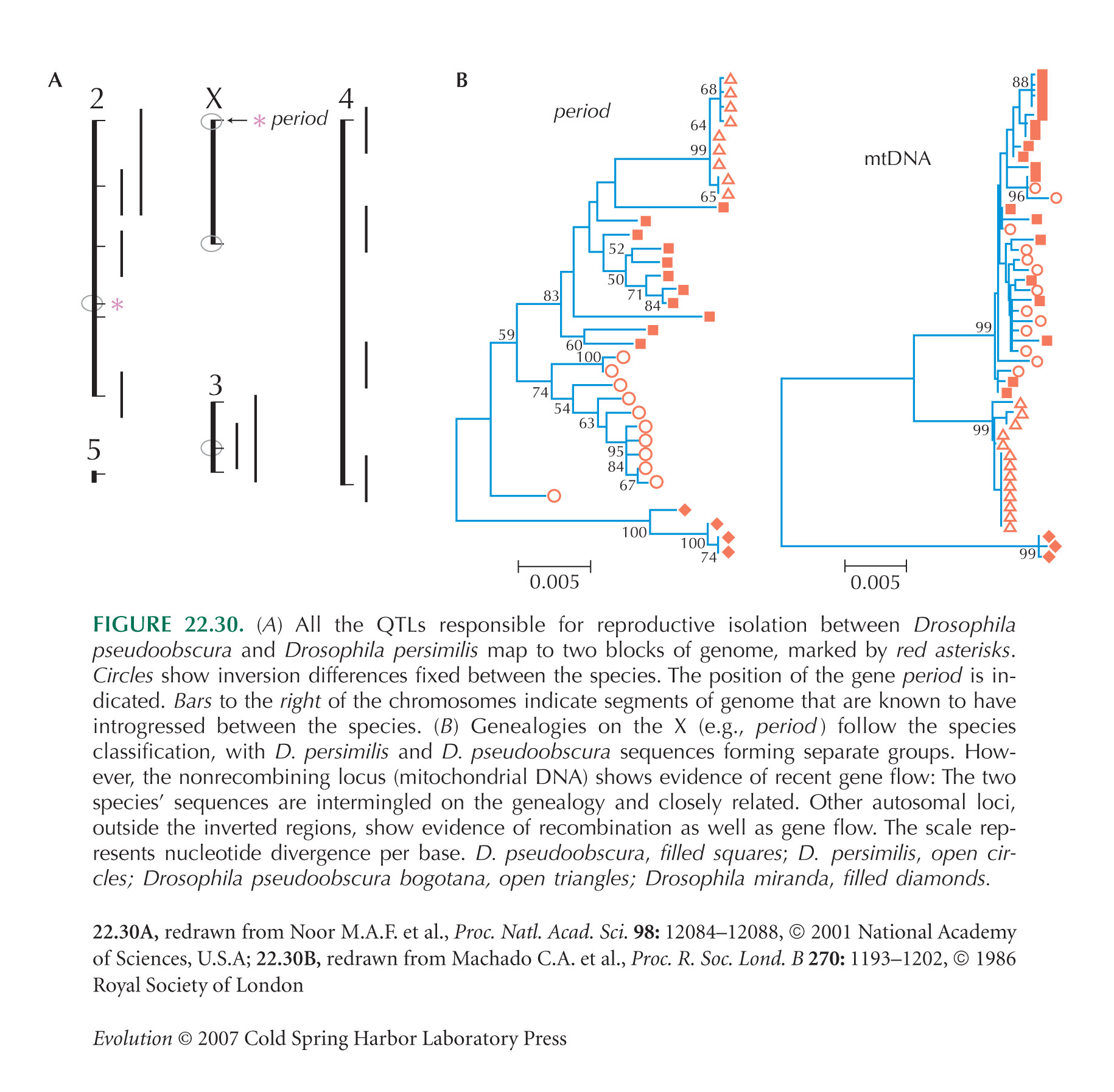
The Drosophila pseudoobscura/persimilis example (Fig. 22.30) is from Noor et al. (2001) and Machado and Hey (2003).
Noor et al. (2001) show striking evidence for the general importance of chromosome rearrangements: Drosophila species pairs that do not differ chromosomally are almost never found in the same place, whereas those with chromosomal differences often are (Table WN22.1; from Noor et al. 2001).
Selection against Crossbreeding Can Reinforce Reproductive Isolation
The idea that reproductive isolation can be strengthened by reinforcement is attractive, because it gives a direct role for natural selection as a cause of speciation. Alfred Russell Wallace (1889) struggled to find a way by which natural selection could favor hybrid sterility, as well as assortative mating. More recently, Dobzhansky (1940) was a strong supporter of the importance of reinforcement and even argued that it could occur on remote islands, driven by selection against interbreeding with occasional long-distance migrants. During the Evolutionary Synthesis, reinforcement came to be seen as the final stage of speciation. Subsequently, there was a reaction against this overenthusiasm (part of a general reaction against “naïve pan-selectionism”; pp. 33–34). Now, however, we have some better evidence as to the actual importance of reinforcement.
It is possible that gene flow between sympatric pairs greatly reduces genetic distance, shifting the points in Figure 22.11A to the left. (We have seen that gene flow has caused substantial mixing of genealogies in the D. pseudoobscura/persimilis example; Fig. 22.30B). However, no difference between sympatric and allopatric populations is evident for postzygotic isolation (Fig. 22.11B), making this objection implausible. Potential difficulties in interpreting Coyne and Orr’s observation of stronger prezygotic isolation between sympatric pairs (Fig. 22.11A) were raised by Kulathinal and Singh (2000).
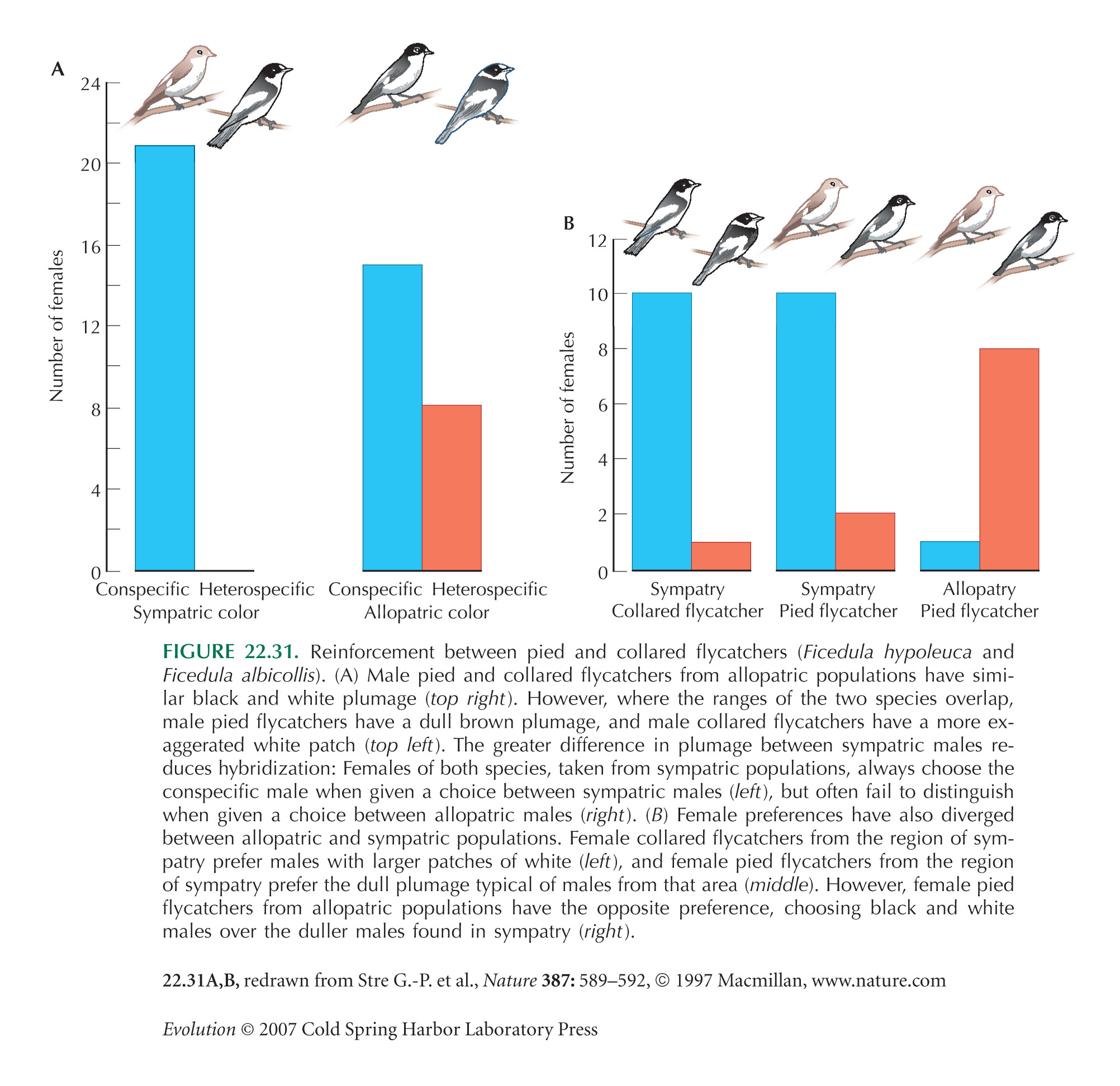
The flycatcher example (Fig. 22.31) is from Saetre et al. (1997); see also Saetre et al. (2001). Veen et al. (2001) have shown that there can be fitness benefits to cross-mating in Ficedula: For example, a male-biased sex ratio in hybrid pairs reduces the costs of producing sterile females (males are fertile).
The Agrostis tenuis flowering time example is from McNeilly and Antonovics (1968). An interesting complication was noted by Stam (1983), who pointed out that differences in flowering time across the boundary would tend to be amplified by differential gene flow.
Selection can strengthen prezygotic isolation through reinforcement, because individuals that avoid mating with the wrong genotype will have fitter offspring. (This can be thought of as a preference for good genes; pp. 581–584.) However, selection cannot strengthen postzygotic isolation, because fitness can only be reduced by a further reduction in the viability or fertility of hybrids. (Darwin appreciated this point in his debates with Wallace; see the first note in this section.) This distinction does not quite hold, however, if siblings compete with each other. In this case, kin selection may favor the elimination of unfit hybrids from a brood of competing siblings, since that will give the other, nonhybrid, siblings an advantage (Coyne, 1974; see also Cronin, 1991, and Problem 22.1).
A Single Population Can Split into Two: Sympatric Speciation
For a view of adaptive radiations, see Schluter (2000).
The phylogenies of the Lake Malawi species flock suggest that divergence has occurred in three stages (Fig. WN22.11). First, there was a separation into rock- and sand-dwelling lineages. (A similar basic split into different ecotypes is seen in sticklebacks.) This was followed by the evolution of different feeding types. A key factor in allowing this diversification was a specialization of the feeding apparatus. The ancestral cichlids had two sets of jaws. The oral jaws collected and processed food, whereas the pharyngeal jaws transported food from the mouth down to the stomach. In the cichlids of the African lakes, the pharyngeal jaws have evolved to process food (cutting prey, stacking scales taken from other fish, and so on), which leaves the oral jaws free to specialize in collecting a wide variety of different foods. Finally, the most closely related species differ primarily in male breeding coloration and are thought to have evolved through sexual selection. There is little postzygotic isolation among cichlid species—fertile hybrids can be produced readily in aquariums—but there is strong prezygotic isolation as hybrids are rarely seen in nature.
The overall timescale of diversification in the cichlids of the three great lakes is comparable with that in the Hawaiian . However, Lake Victoria dried out completely only 15,000 years ago. It had been thought that the hundreds of species now found there had diversified within this extremely short time, an argument supported by the first studies of mtDNA, which showed the Lake Victoria species to be monophyletic. However, recent work has sampled more widely around Lake Victoria, more variable regions of the mtDNA, and multiple markers across the nuclear genome. This has made it clear that many cichlid species survived the desiccation, so that much of the diversity seen in Lake Victoria now dates from well before the local extinction. Although the “superflock” of cichlids in the Lake Victoria region (including Lakes Kivu, Edward, and George) is monophyletic, the cichlids within Lake Victoria are not. Mitochondrial diversity is extremely low within the “superflock.” However, nuclear gene diversity is much higher, and so this homogeneity is due either to chance or to the introgression of mtDNA across species boundaries (see above), rather than to very recent diversification of the species.
Danley and Kocher (2001) provide a good overview of recent work on the great lake cichlids (although before Verheyen et al. [2003] and Seehausen et al. [2003] showed that the Lake Victoria radiation is not so very recent). Kornfield and Smith (2000) is also a good recent review of the system, and Goldschmidt (1998) provides a nice popular account. This Web site contains abundant information about the Lake Malawi cichlids and an extensive bibliography.
The Cameroon crater lake example (Fig. 22.33) is from Schliewen et al. (1994, 2001). Schliewen and Klee (2004) give further evidence for the monophyletic origin of these taxa.
For other plausible examples of symptaric speciation, see Savolainen et al. (2006) and Barluenga et al. (2006), but also see criticisms by Stuessy (2006) and by Schliewen et al. (2006).
Maynard Smith (1966) provides an influential early model of sympatric speciation. Felsenstein (1981) first emphasized the important distinction between models where one allele or trait must be established to produce assortative mating between differently adapted types, as opposed to those where different alleles must become associated with the alternative types, in linkage disequilibrium. Coyne and Orr (2004, Chapter 4) and Bolnick and Fitzpatrick (2007) give critical reviews of the recent enthusiasm for sympatric speciation.
Dieckmann and Doebeli (1999) introduced an influential model of sympatric speciation, in which disruptive selection arises spontaneously through specialization of competing genotypes within an initially uniform distribution of resources. (The basic model was first introduced by Roughgarden [1972].) Difficulties with the Dieckmann and Doebeli model are discussed by Gavrilets (2004, Chapter 10) and by Polechova and Barton (2005). For an interesting survey of its influence, see Butlin and Tregenza (2005).
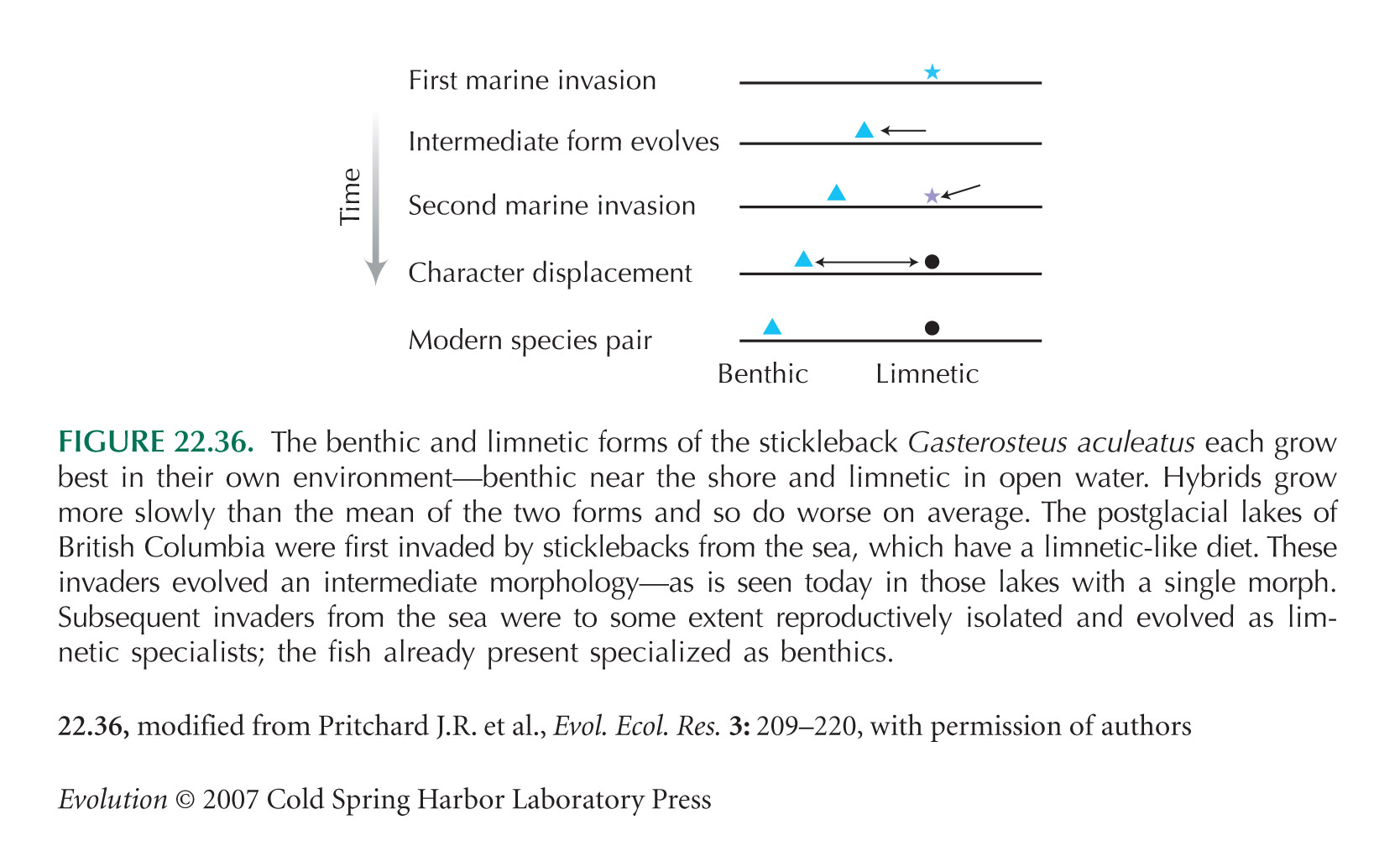
The mitochondrial DNA phylogeny of sticklebacks from British Columbia suggests that the forms within each lake are monophyletic, which would be consistent with sympatric speciation within each lake. However, nuclear markers support the double-invasion hypothesis described in the text. Flow of mitochondrial genomes between feeding morphs makes the mtDNA genealogy misleading. This is another example of the pattern noted above. See Schluter (1996).
The stickleback data shown in Figure 22.36 are from Schluter (1996) and Pritchard and Schluter (2001). Figures 2 and 4 of Schluter (1996) give evidence that the benthic and limnetic forms are adapted to their different environments and that hybrids are less fit in either.
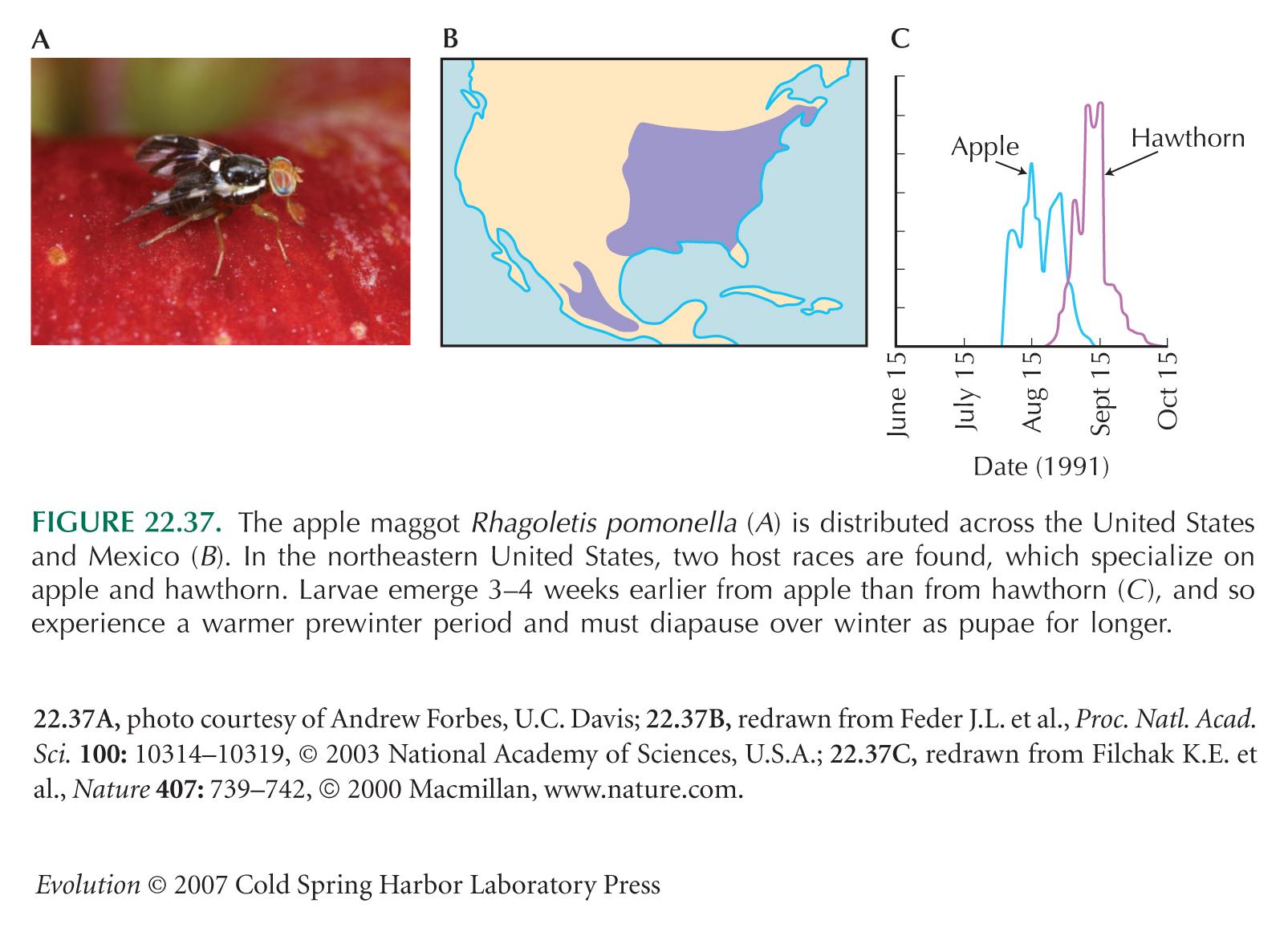
Host Races Provide a Classic Example of Incipient Speciation
Divergence of host races in Rhagoletis was described by Walsh (1864, 1867), who also proposed that such host races could lead to sympatric speciation.
The Rhagoletis example of Figure 22.37 is from Feder et al. (2003) and Filchak et al. (2000).
Filchak et al. (2000) show intriguing evidence of strong selection on allozyme markers: When larvae are reared at warm temperatures before winter, those carrying alleles from the hawthorn race survive more poorly when exposed to long overwinter periods—a regime typical of apples. However, this pattern is reversed when prediapause temperatures are low. There is a complex three-way interaction between per-winter temperature, overwinter length, and genotype, whose net effect is to favor apple alleles in an apple-like environment and vice versa.
References
Barbash D.A., Roote J., and Ashburner M. 2000. The Drosophila melanogaster hybrid male rescue gene causes inviability in male and female species hybrids. Genetics 154: 1747–1771.
Barluenga M., Stolting K.N., Salzburger W., Muschick M., and Meyer A. 2006. Sympatric speciation in Nicaraguan crater lake cichlid fish. Nature 439: 719–723 and 444: E12–E13.
Barraclough T.G. and Vogler A.P. 2000. Detecting the geographical pattern of speciation from species-level phylogenies. Am. Nat. 155: 419–434.
Barraclough T.G., Birky C.W., and Burt A. 2003. Diversification in sexual and asexual organisms. Evolution 57: 2166–2172.
Barrett S.J. and Sneath P.H.A. 1994. A numerical phenotypic taxonomic study of the genus Neisseria. Microbiology 140: 2867–2891.
Barton N.H. 1989. Founder effect speciation. In Speciation and its consequences (ed. D. Otte and J.A. Endler), Chap. 10. Sinauer Press, Sunderland, Massachusetts.
Barton N.H. 1998. Natural selection and random genetic drift as causes of evolution on islands In Evolution on islands (ed. P.R. Grant), pp. 102–123. Oxford University Press, Oxford.
Barton N.H. and Charlesworth B. 1984. Genetic revolutions, founder effects, and speciation. Annu. Rev. Ecol. Syst. 15: 133–164.
Bateson W. 1909. Heredity and variation in modern lights. In Darwin and modern science (ed. A. C. Seward), pp. 85–101. Cambridge University Press, Cambridge.
Berlin B. 1973. Folk systematics in relation to biological classification and nomenclature. Annu. Rev. Ecol. Syst. 4: 259–271.
Bolnick D.I. and Fitzpatrick B.M. 2007. Sympatric speciation: Models and empirical evidence. Annu. Rev. Ecol. Syst. 38: 459–487.
Bradshaw H.D., Wilbert S.M., Otto K.G., and Schemske D.W. 1995. Genetic mapping of floral traits associated with reproductive isolation in monkeyflowers (Mimulus). Nature 376: 762–765.
Bradshaw H.D., Otto K.G., Frewen B.E., McKay J.K., and Schemske D.W. 1998. Quantitative trait loci affecting differences in floral morphology between two species of monkeyflower (Mimulus). Genetics 149: 367–382.
Buerkle C.A. and Rieseberg L.H. 2001. Low intraspecific variation for genomic isolation between hybridizing sunflower species. Evolution 55: 684–691.
Bush G.L., Case S.M., Wilson A.C., and Patton J.L. 1977. Rapid speciation and chromosomal evolution in mammals. Proc. Natl. Acad. Sci. 74: 3942–3946.
Butlin R. and Tregenza T. 2005. The way the world might be. J. Evol. Biol. 18: 1205–1208.
Carson H.L. 1987. Tracing ancestry with chromosomal sequences. Trends Ecol. Evol. 2: 203–207.
Carson H.L. and Kaneshiro K.Y. 1976. Drosophila of Hawaii: Systematics and ecological genetics. Annu. Rev. Ecol. Syst. 7: 311–346.
Carson H.L. and Templeton A.R. 1984. Genetic revolutions in relation to speciation phenomena: The founding of new populations. Annu. Rev. Ecol. Syst. 15: 97–131.
Charlesworth B., Coyne J.A., and Barton N.H. 1987. The relative rates of evolution of sex chromosomes and autosomes. Am. Nat. 129: 113–146.
Cohan F.M. 2002. What are bacterial species? Annu. Rev. Microbiol. 56: 457–487.
Coyne J.A. 1974. The evolutionary origin of hybrid inviability. Evolution 28: 505–506.
Coyne J.A. 1985. The genetic basis of Haldane’s Rule. Nature 314: 736–738.
Coyne J.A. and Orr H.A. 1997. Patterns of speciation in Drosophila revisited. Evolution 51: 295–303.
Coyne J.A. and Orr H.A. 2004. Speciation. Sinauer Press, Sunderland, Massachusetts.
Craddock E.M. 2000. Speciation processes in the adaptive radiation of Hawaiian plants and animals. Evol. Biol. 31: 1–53.
Cronin H. 1991. The ant and the peacock: Altruism and sexual selection from Darwin to today. Cambridge University Press, Cambridge.
Danley P.D. and Kocher T.D. 2001. Speciation in rapidly diverging systems: Lessons from Lake Malawi. Mol. Ecol. 10: 1075–1086.
Desmond A. and Moore J.R. 1992. Darwin. Michael Joseph, London.
Dieckmann U. and Doebeli M. 1999. On the origin of species by sympatric speciation. Nature 400: 354–357.
Dobzhansky T. 1936. Studies on hybrid sterility. II. Localization of sterility factors in Drosophila pseudoobscura hybrids. Genetics 21: 113–135.
Dobzhansky T. 1940. Speciation as a stage in evolutionary divergence. Am. Nat. 74: 312–321.
Endler J.A. 1977. Geographic variation, speciation, and clines. Princeton University Press, Princeton, New Jersey.
Endler J.A. 1982. Problems in distinguishing historical from ecological factors in biogeography. Am. Zool. 22: 441–452.
Feder J.L., Berlocher S.H., Roethele J.B., Dambroski H., Smith J.J., et al. 2003. Allopatric genetic origins for sympatric host-plant shifts and race formation in Rhagoletis. Proc. Natl. Acad. Sci. 100: 10314–10319.
Felsenstein J. 1975. A pain in the torus: Some difficulties with the model of isolation by distance. Am. Nat. 109: 359–368.
Felsenstein J. 1981. Skepticism towards Santa Rosalia, or why are there so few kinds of animals? Evolution 35: 124–138.
Filchak K.E., Roethele J. B., and Feder J.L. 2000. Natural selection and sympatric divergence in the apple maggot Rhagoletis pomonella. Nature 407: 739–742.
Fitzpatrick B.M. and Turelli M. 2006. The geography of mammalian speciation: Mixed signals from phylogenies and range maps. Evolution 60: 601–615.
Fontaneto D., Herniou E.A., Boschetti C., Caprioli M., Melone G., Ricci C., and Barraclough T.G. 2007. Independently evolving species in asexual bdelloid rotifers. PLoS Biol. 5: 87–95.
Gao L.Z. and Zhang J.Z. 2003. Why are some human disease-associated mutations fixed in mice? Trends Genet. 19: 678–681.
Gavrilets S. 2004. Fitness landscapes and the origin of species. Princeton University Press, Princeton, New Jersey.
Giddings L.V., Kaneshiro K.Y., and Anderson W.W. 1989. Genetics, speciation and the founder principle. Oxford University Press, Oxford.
Gevers D., Cohan F.M., Lawrence J.G., Spratt B.G., Coenye T., et al. 2005. Re-evaluating prokaryotic species. Nat. Rev. Microbiol. 3: 733–739.
Goldschmidt T. 1998. Darwin’s dreampond: Drama on Lake Victoria. MIT Press, Cambridge, Massachusetts.
Gould S.J. 1980. A quahog is a quahog. In The panda’s thumb: More reflections in natural history, 1st ed., pp. 204–213. W.W. Norton, New York.
Haldane J.B.S. 1922. Sex ratio and unisexual sterility in hybrid animals. J. Genet. 12: 101–109.
Harrison R.G. 1998. Linking evolutionary pattern and process: The relevance of species concepts for the study of speciation. In Endless forms: Species and speciation (ed. D.J. Howard and S.H. Berlocher), pp. 19–31. Oxford University Press, New York.
Heliovaara K., Vaisanen R., and Simon C. 1994. Evolutionary ecology of periodical insects. Trends Ecol. Evol. 9: 475–481.
Holder M.T., Erdmann M.V., Wilcox T.P., Caldwell R.L., and Hillis D.M. 1999. Two living species of coelacanths? Proc. Natl. Acad. Sci. 96: 12616–12620.
Holman E.W. 1987. Recognizability of sexual and asexual species of rotifers. Syst. Zool. 36: 381–386.
Howard D.J., Preszler R.W., Williams J., Fenchel S., and Boecklen W.J. 1997. How discrete are oak species? Insights from a hybrid zone between Quercus grisea and Quercus gambelii. Evolution 51: 747–755.
Hubbell S. 2001. The unified neutral theory of biodiversity and biogeography. Princeton University Press, Princeton, New Jersey.
Hudson R.R. and Coyne J. 2002. Mathematical consequences of the genealogical species concept. Evolution 56: 1557–1565.
Hughes A.L. 1999. Adaptive evolution of genes and genomes. Oxford University Press, Oxford.
Irwin D.E., Irwin J.H., and Price T.D. 2001. Ring species as bridges between microevolution and speciation. Genetica 112–113: 223–243.
Katinka M.D., Duprat S., Cornillot E., Metenier G., Thomarat F., et al. 2001. Genome sequence and gene compaction of the eukaryote parasite Encephalitozoon cuniculi. Nature 414: 450–453.
Keeling P.J. 2001. Parasites go the full monty. Nature 414: 401–402.
Kern A.D. and Kondrashov F.A. 2004. Mechanisms and convergence of compensatory evolution in mammalian mitochondrial tRNAs. Nat. Genet. 36: 1207–1212. Epub 2004 Oct 24.
Klicka J. and Zink R.M. 1999. Pleistocene effects on North American songbird evolution. Proc. R. Soc. Lond. B 266: 695–700.
Kondrashov A.S., Sunyaev S., and Kondrashov F.A. 2002. Dobzhansky–Muller incompatibilities in protein evolution. Proc. Natl. Acad. Sci. 99: 14878–14883.
Kornfield I. and Smith P.F. 2000. African cichlid fishes: Model systems for evolutionary biology. Annu. Rev. Ecol. Syst. 31: 163–196.
Kulathinal R., Bettencourt B.R., and Hartl D. 2004. Compensated deleterious mutations in insect genomes. Science 306: 1553–1555.
Kulathinal R.J. and Singh R.S. 2000. Reinforcement with gene flow? A reply. Evolution 54: 2176–2177.
Lande R. 1979. Effective deme sizes during long-term evolution estimated from rates of chromosomal rearrangement. Evolution 33: 234–251.
Lande R. 1981. The minimum number of genes contributing to quantitative variation between and within populations. Genetics 99: 541–553.
Lande R. 1984. The expected fixation rate of chromosomal inversions. Evolution 38: 743–752.
Lande R. 1985. The fixation of chromosomal rearrangements in a subdivided population with local extinction and recolonisation. Heredity 54: 323–332.
Levin D.A. and Wilson A.C. 1976. Rates of evolution in seed plants: Net increase in diversity of chromosome numbers and species numbers through time. Proc. Natl. Acad. Sci. 73: 2086–2090.
Lloyd J.E. 1966. Studies on the flash communication system in Photinus fireflies. Misc. Publ. Univ. Mus. Zool. Univ. Michigan 130: 1–195.
Love A., Love D., and Pichi Sermolli R.E.G. 1977. Cytotaxonomical atlas of the Pteridophyta. Vaduz, Germany.
Lovette I.J. 2005. Glacial cycles and the tempo of avian speciation. Trends Ecol. Evol. 20: 57–59.
Lynch M. 2007. The origins of genome architecture. Sinauer Associates, Sunderland, Massachussetts.
Machado C.A. and Hey J. 2003. The causes of phylogenetic conflict in a classic Drosophila species group. Proc. R. Soc. Lond. B 270: 1193–1202.
Macholan M., Munclinger P., Ugerkova M., Dufkova P., Bimová B., Bolková E., Zima J., and Pialek J. 2007. Genetic analysis of autosomal and X-linked markers across a mouse hybrid zone. Evolution 61: 746–771.
McNeilly T. and Antonovics A. 1968. Evolution in closely adjacent populations. IV. Barriers to gene flow. Heredity 23: 205–218.
Masly J.P. and Presgraves D.C.. 2007. High-resolution genome-wide dissection of the two rules of speciation in Drosophila. PLoS Biol. 5(9): e243. Published online 2007 September 11. doi:10.1371/journal.pbio.0050243.
Maynard Smith J. 1966. Sympatric speciation. Am. Nat. 100: 637–650.
Maynard Smith J. and Szathmáry E. 1995. The major transitions in evolution. W.H. Freeman, Oxford.
Mayr E. 1963. Animal species and evolution. Harvard University Press, Cambridge, Massachusetts.
Mayr E. and O’Hara R.J. 1986. The biogeographic evidence supporting the Pleistocene forest refuge hypothesis. Evolution 40: 55–67.
Mendelson T.C. 2003. Sexual isolation evolves faster than hybrid inviability in a diverse and sexually dimorphic genus of fish (Percidae: Etheostoma). Evolution 57: 317–327.
Muller H.J. 1925. Why polyploidy is rarer in animals than plants. Am. Nat. 59: 346–353.
Muller H.J. 1940. Bearings of the Drosophila work on systematics. In The new systematics (ed. J. Huxley), pp. 185–268. Clarendon Press, Oxford.
Muller H.J. 1942. Isolating mechanisms, evolution and temperature. Biol. Symp. 6: 71–125.
Müntzing A. 1930. Outlines to a genetic monograph of the genus Grrleopsis. Hereditas 13: 185–341.
Murray J. and Clarke B.C. 1980. The genus Partula: Speciation in progress. Proc. R. Soc. Lond. B 211: 83–117.
Navarro A. and Barton N.H. 2003. Accumulating postzygotic isolation genes in parapatry: A new twist on chromosomal speciation. Evolution 57: 447–459.
Noor M.A.F., Grams K.L., Bertucci L.A., and Reiland J. 2001. Chromosomal inversions and the reproductive isolation of species. Proc. Natl. Acad. Sci. 98: 12084–12088.
Orr H.A. 1990. Why polyploidy is rarer in animals than in plants revisited. Am. Nat. 136: 759–770.
Orr H.A. 1997a. Haldane’s Rule. Annu. Rev. Ecol. Syst. 28: 195–218.
Orr H.A. 1997b. Dobzhansky, Bateson and the genetics of speciation. Genetics 144: 1331–1335.
Orr H.A. 2001. The genetics of species differences. Trends Ecol. Evol. 16: 343–358.
Orr H.A. 2005. The genetic basis of reproductive isolation: Insights from Drosophila. Proc. Natl. Acad. Sci. 102: 6522–6526.
Orr H.A. and Coyne J.A. 1992. The genetics of adaptation: A reassessment. Am. Nat. 140: 725–742.
Orr H.A. and Irving S. 2000. Genetic analysis of the hybrid male rescue locus of Drosophila. Genetics 155: 225–231.
Orr H.A. and Irving S. 2001. Complex epistasis and the genetic basis of hybrid sterility in the Drosophila pseudoobscura Bogota-USA hybridization. Genetics 158: 1089–1100.
Orr H.A. and Irving S. 2005. Segregation distortion in hybrids between the Bogota and USA subspecies of Drosophila pseudoobscura. Genetics 169: 671–682.
Orr H.A. and Presgraves D.C. 2000. Speciation by postzygotic isolation: Forces, genes and molecules. BioEssays 22: 1085–1094.
Orr H.A., Masly J.P., and Presgraves D.C. 2004. Speciation genes. Curr. Opin. Genet. Dev. 14: 675–679.
Otto S.P. and Whitton J. 2000. Polyploid incidence and evolution. Annu. Rev. Genet. 34: 401–437.
Payseur B.A., Krenz J.G., and Nachman M.W. 2004. Differential patterns of introgression across the X chromosome in a hybrid zone between two species of house mice. Evolution 58: 2064–2078.
Polechova J. and Barton N.H. 2005. Speciation through competition: A critical review. Evolution 59: 1194–1210.
Pounds J.A. and Jackson J.F. 1981. Riverine barriers to gene flow and the differentiation of fence lizard populations. Evolution 35: 516–528.
Poulton E.B. 1908. What is a species? In Essays on evolution: 1889–1907, pp. 46–94. Oxford University Press, Oxford.
Presgraves D.C. 2002. Patterns of postzygotic isolation in Lepidoptera. Evolution 56: 1168–1183.
Presgraves D.C. 2003. A fine-scale genetic analysis of hybrid incompatibilities in Drosophila. Genetics 163: 955–972.
Presgraves D.C. and Orr H.A. 1998. Haldane’s Rule in taxa lacking a hemizygous X. Science 282: 952–954.
Presgraves D.C. and Stephan W. 2007. Pervasive adaptive evolution among interactors of the Drosophila hybrid inviability gene, Nup96. Mol. Biol. Evol. 24: 306–314.
Presgraves D.C., Balagopalan L., Abmayr S.M., and Orr H.A. 2003. Adaptive evolution drives divergence of a hybrid inviability gene between two species of Drosophila. Nature 423: 715–719.
Price T.D. and Bouvier M.M. 2002. The evolution of F1 postzygotic incompatibilities in birds. Evolution 56: 2083–2089.
Pritchard J.R. and Schluter D. 2001. Declining interspecific competition during character displacement: Summoning the ghost of competition past. Evol. Ecol. Res. 3: 209–220.
Ramsey J., Bradshaw H.D., and Schemske D.W. 2003. Components of reproductive isolation between Mimulus lewisii and M. cardinalis (Scrophulariaceae). Evolution 57: 1520–1534.
Ramsing N.B., Ferris M.J., and Ward D.M. 2000. Highly ordered vertical structure of Synechococcus populations within the one-millimetre thick photic zone of a hot spring cyanobacterial mat. Appl. Environ. Microbiol. 66: 1038–1049.
Read A. and Nee S. 1991. Is Haldane’s Rule significant ? Evolution 45: 1707–1710.
Rice W.R. and Hostert E.E. 1993. Laboratory experiments on speciation: What have we learned in forty years ? Evolution 47: 1637–1653.
Rieseberg L.H. 1997. Hybrid origins of plant species. Annu. Rev. Ecol. Syst. 28: 359–389.
Rieseberg L.H. and Noyes R.D. 1998. Genetic map-based studies of reticulate evolution in plants. Trends Plant Sci. 3: 254–259.
Rieseberg L.H. and Soltis D.E. 1991. Phylogenetic consequences of cytoplasmic gene flow in plants. Evol. Trends Plants 5: 65–84.
Rieseberg L.H., Linder C.R., and Seiler G.J. 1995a. Chromosomal and genic barriers to introgression in Helianthus. Genetics 141: 1163–1171.
Rieseberg L.H., Van Fossen C., and Descrochers A.M. 1995b. Genomic reorganisation accompanies hybrid speciation in wild sunflowers. Nature 375: 313–316.
Rieseberg L.H., Sinervo B., Linder C.R., Ungerer M.C., and Arias D.M. 1996. Role of genetic interactions in hybrid speciation: Evidence from ancient and experimental hybrids. Science 272: 741–745.
Roberts M.S. and Cohan F.M. 1993. The effect of DNA sequence divergence on sexual isolation in Bacillus. Genetics 134: 401–408.
Rogers C.E., Thompson T.E., and Seiler G.J. 1982. Sunflower species of the United States. National Sunflower Association, Bismarck, North Dakota.
Roughgarden J. 1972. The evolution of niche width. Am. Nat. 106: 683–718.
Russell S.T. 2003. Evolution of intrinsic post-zygotic reproductive isolation in fish. Ann. Zool. Fenn. 40: 321–329.
Saetre G.-P., Borge T., Lindell J., Moum T., Primmer C.R., et al. 2001. Speciation, introgressive hybridization and nonlinear rate of molecular evolution in flycatchers. Mol. Ecol. 10: 737–750.
Saetre G.-P., Moum T., Bures S., Král M., Adamjan M., and Moreno T. 1997. A sexually selected character displacement in flycatchers reinforces premating isolation. Nature 387: 589–592.
Savolainen V., Anstett M.C., Lexer C., Hutton I., Clarkson J.J., Norup M.V., Powell M.P., Springate D., Salamin N., and Baker W.J. 2006. Sympatric speciation in palms on an oceanic island. Nature 441: 210–213, 443: E12–E13.
Sawamura K., Taira T., and Watanabe T.K. 1993a. Hybrid lethal systems in the D. melanogaster species complex. I. The maternal hybrid rescue (mhr) gene of D. simulans. Genetics 133: 299–305.
Sawamura K., Yamamoto M.T., and Watanabe T.K. 1993b. Hybrid lethal systems in the D. melanogaster species complex. II. The zygotic hybrid rescue (zhr) gene of D. melanogaster. Genetics 133: 307–313.
Schliewen U.K. and Klee B. 2004. Reticulate sympatric speciation in Cameroonian crater lake cichlids. Front. Zool. 1: 5.
Schliewen U.K., Tautz D., and Paabo S. 1994. Sympatric speciation suggested by monophyly of crater lake cichlids. Nature 368: 629–632.
Schliewen U.K., Rassmann K., Markmann M., Markert J.A., Kocher T., et al. 2001. Genetic and ecological divergence of a monophyletic cichlid species pair under fully sympatric conditions in Lake Ejagham, Cameroon. Mol. Ecol. 10: 1471–1488.
Schliewen U.K., Kocher T.D., McKaye K.R., Seehausen O., and Tautz D. 2006. Evidence for sympatric speciation? Nature 444: E12–E13.
Schluter D. 1996. Ecological causes of adaptive radiation. Am. Nat. 148: S40–S64.
Schluter D. 2000. The ecology of adaptive radiation. Oxford University Press, Oxford.
Schwarzbach A.E., Donovan L.A., and Rieseberg L.H. 2001. Transgressive character expression in a hybrid sunflower species. Am. J. Bot. 88: 270–277.
Searle J.B. 1986. Factors responsible for a karyotypic polymorphism in the common shrew, Sorex araneus. Proc. R. Soc. Lond. B 229: 277–298.
Searle J.B. 1993. Chromosomal hybrid zones in eutherian mammals. In Hybrid zones and the evolutionary process (ed. R.G. Harrison), pp. 309–354. Oxford University Press, Oxford.
Seehausen O., Koetsier E., Schneider M.V., Chapman L.J., Chapman C.A., et al. 2003. Nuclear markers reveal unexpected genetic variation and a Congolese-Nilotic origin of the Lake Victoria cichlid species flock. Proc. R. Soc. Lond. B 270: 129–137.
Seoighe C. 2003. Turning the clock back on ancient genome duplication. Curr. Opin. Genet. Dev. 13: 636–643.
Sepp S. and Paal J. 1998. Taxonomic continuum of Alchemilla (Rosaceae) in Estonia. Nord. J. Bot. 18: 519–535.
Shuker D.M., Underwood K., King T.M., and Butlin R.K. 2005. Patterns of male sterility in a grasshopper hybrid zone imply accumulation of hybrid incompatibilities without selection. Proc. Biol. Sci. 272: 2491–2497.
Stam P. 1983. The evolution of reproductive isolation in closely adjacent plant populations through differential flowering time. Heredity 50: 105–118.
Stuessy T.F. 2006. Sympatric plant speciation on islands? Nature 443: E12.
Sucena E. and Stern D. 2000. Divergence of larval morphology between Drosophila sechellia and its sibling species caused by cis-regulatory evolution of ovo/shaven-baby. Proc. Natl. Acad. Sci. 97: 4530–4534.
Sucena E., Delon I., Jones I.L., Payre F., and Stern D.L. 2003. Regulatory evolution of shavenbaby/ovo underlies multiple cases of morphological parallelism. Nature 424: 935–938.
Tamura K., Subramaniam S., and Kumar S. 2004. Temporal patterns of fruit fly (Drosophila) evolution revealed by mutation clocks. Mol. Biol. Evol. 21: 36–44.
Templeton A.R. 1977. Analysis of head shape differences between two interfertile species of Hawaiian Drosophila. Evolution 31: 630–642.
True J.R., Weir B.S., and Laurie C.C. 1996. A genome wide survey of hybrid incompatibility factors by the introgression of marked segments of Drosophila mauritiana chromosomes into Drosophila simulans. Genetics 142: 819–837.
Turelli M. and Begun D.J. 1997. Haldane’s rule and X-chromosome size in Drosophila. Genetics 147: 1799–1815.
Uhl C.H. 1978. Chromosomes of Mexican sedum. II. Section Pachysedum. Rhodora 80: 491–512.
Ungerer M.C., Baird S.J.E., Pan J., and Rieseberg L.H. 1998. Rapid hybrid speciation in wild sunflowers. Proc. Natl. Acad. Sci. 95: 11757–11762.
Vanlerberghe F., Boursot P., Nielsen J.T., and Bonhomme F. 1988. A steep cline for mitochondrial DNA in Danish mice. Genet. Res. 52: 185–193.
Vanlerberghe F., Dod B., Bellis M., and Bonhomme F. 1986. Absence of Y-chromosome introgression across the hybrid zone between Mus musculus domesticus and Mus musculus musculus. Genet. Res. 48: 191–197.
Veen T., Borge T., Griffith S.C., Saetre G.P., Bures S., et al. 2001. Hybridization and adaptive mate choice in flycatchers. Nature 411: 45–50.
Verheyen E., Salzburger W., Snoeks J., and Meyer A. 2003. Origin of the superflock of cichlid fishes from Lake Victoria, East Africa. Science 300: 325–329.
Vincek V., O’Huigin C., Satta Y., et al. 1997. How large was the founding population of Darwin’s finches? Proc. R. Soc. Lond. B 264: 111–118.
Virdee S.R. and Hewitt G.M. 1992. Postzygotic isolation and Haldane’s Rule in a grasshopper. Heredity 69: 527–538.
Wallace A.R. 1889. Darwinism. Macmillan, London.
Walsh B.D. 1864. On phytophagic varieties and phytophagic species. Proc. Entomol. Soc. Philadelphia 3: 403–430.
Walsh B.D. 1867. The apple-worm and the apple maggot. J. Hortic. 2: 338–343.
Walsh J.B. 1982. Rate of accumulation of reproductive isolation by chromosome rearrangements. Am. Nat. 120: 510–532.
Whittemore A.T. and Schaal B.A. 1991. Interspecific gene flow in sympatric oaks. Proc. Natl. Acad. Sci. 88: 2540–2544.
Wilson A.C., Bush G.L., Case S.M., and King M.C. 1975. Social structuring of mammalian populations and rate of chromosomal evolution. Proc. Natl. Acad. Sci. 72: 5061–5065.
Wolfe K.H. 2001. Yesterday’s polyploids and the mystery of diploidization. Nat. Rev. Genet. 2: 333–341.
Wright S. 1941. On the probability of fixation of reciprocal translocations. Am. Nat. 75: 513–522.
Young W.R., Roberts A.J., and Stuhne G. 2001. Reproductive pair correlations and the clustering of organisms. Nature 412: 328–331.
Zeng Z.B., Liu J.J., Stam L.F., Kao C.H., Mercer J.M., et al. 2000. Genetic architecture of a morphological shape difference between two Drosophila species. Genetics 154: 299–310.
|



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}