Chapter 23 Notes
Evolution of Genetic Systems
Introduction
The opening figure (p. 657) shows a human egg and sperm.
Studying the Evolution of Genetic Systems
All Aspects of Genetic Systems Evolve
Experimental evidence that there is heritable variation in recombination rates is reviewed by Brooks (1988); there is variation both in the overall rate and in rates at specific sites in the genome. Coop et al. (2008) estimate recombination rates from human pedigrees, using large numbers of SNP markers, and also find heritable variation.
The Evolution of Genetic Systems Can Be Studied in Several Ways
The evolution of genetic systems can be understood from the fate of alleles that modify them. We discuss this approach, and its history, .
Evolution of Mutation Rates
The Cost of Preventing Harmful Mutations Drives the Evolution of Mutation Rates
Nothel (1987) selected for reduced mutation rates in Drosophila. Schaaper (1998) reviews work on antimutators. Sniegowski et al. (2000) give a brief summary.
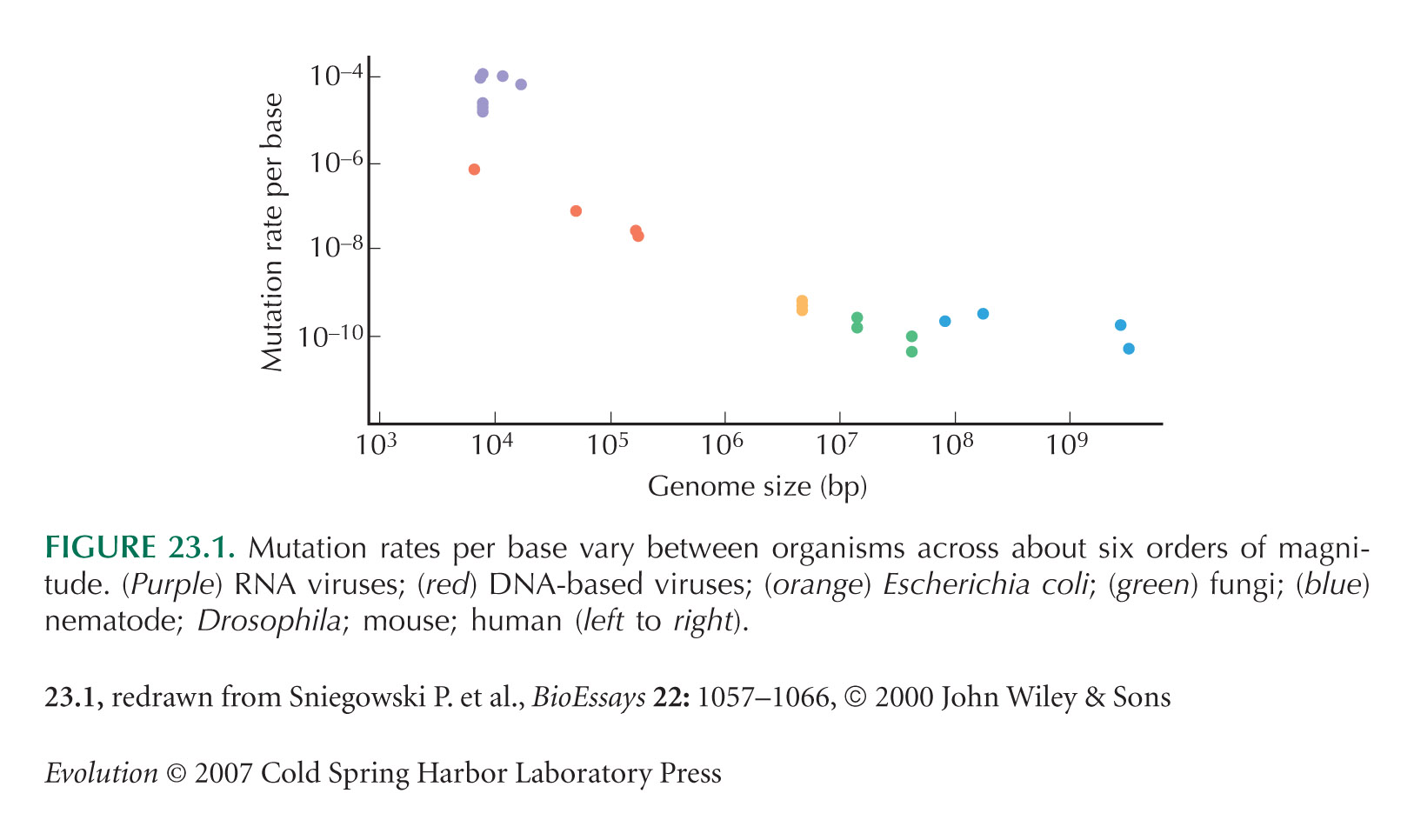
Drake et al. (1998) and Drake and Holland (1999) discuss mutation load in HIV and other RNA viruses. Figure 23.1 is from Sniegowski et al. (2000), based on data from Drake et al. (1998).
Dawson (1998) sets out the theoretical balance between the cost of reducing mutation and the mutation load.
Mutator Alleles Can Hitchhike to High Frequency with the Beneficial Alleles That They Cause
Matic et al. (1997) survey in natural populations of Escherichia coli.
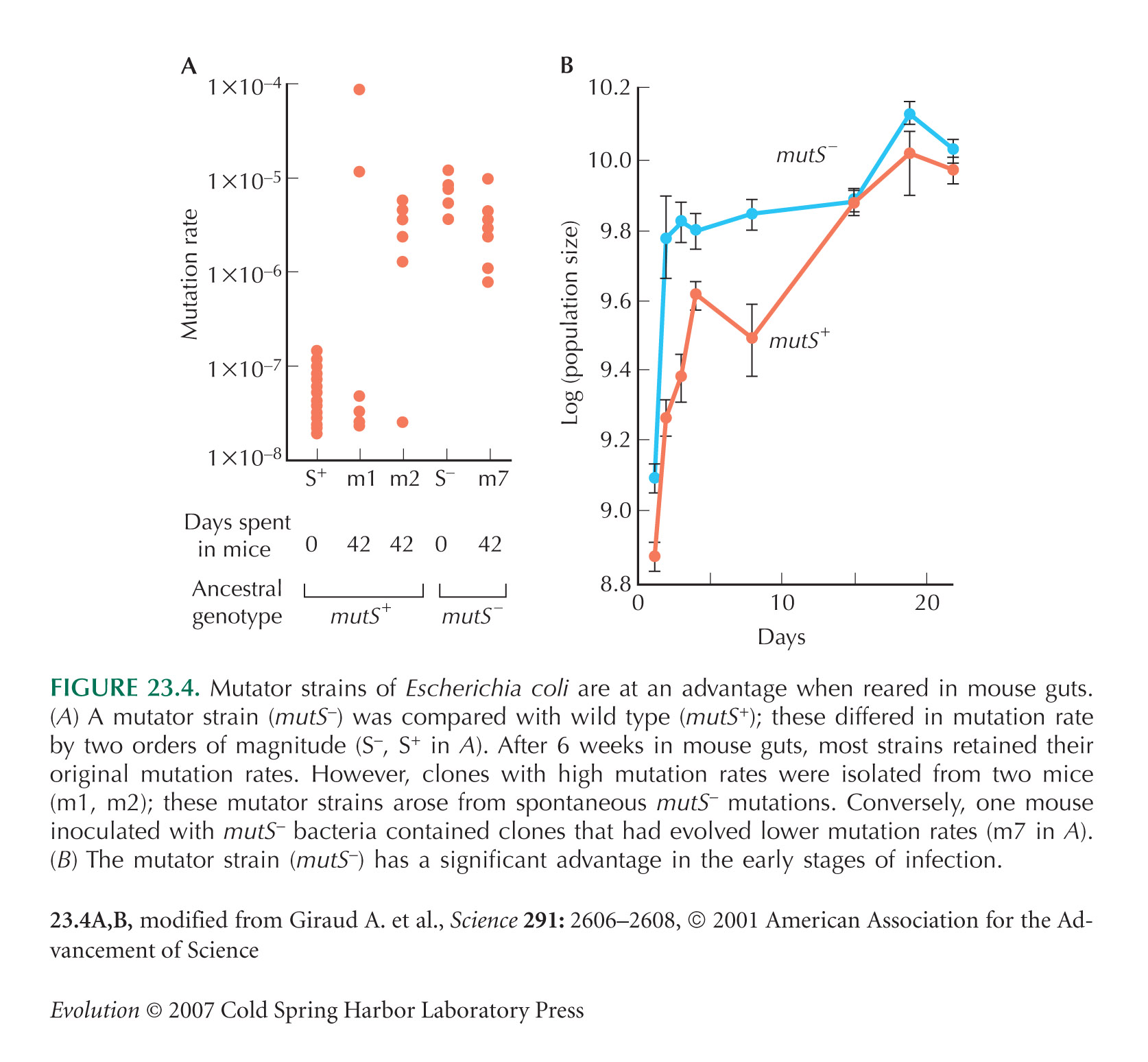
The example of Figure 23.4 is from Giraud et al. (2001).
Some Genes Have Evolved Mechanisms to Raise Their Mutation Rate
Note that both at contingency loci and in the vertebrate immune system, variation is generated by recombination at the molecular level. However, this recombination is between repeated sequences within the same genome. Their population genetic effect is therefore to produce random mutation, which is why we discuss these examples here.
Tomlinson et al. (1996) review evidence on mutation rates in cancer cells. They conclude that normal mutation rates are sufficient to explain observed rates of cancer. Elevated mutation rates are often found, but it is not clear whether these evolved early or late in tumor progression.
Long-term selection experiments by Sniegowski et al. (1997) showed mismatch repair mutations causing higher mutation rates in 3 of 12 lines.
We detail examples of contingency loci .
Although all vertebrates share the same basic immune system, there is considerable variation in just how genetic diversity is generated. For example, chickens have only one functional set of the VL, JL and VH, DH, JH regions, but they carry large numbers of degenerate copies. Gene conversion from these to the functional copies generates variation, in much the same way as the pilE example in Neisseria meningitidis discussed .
Evolution of Sex and Recombination
Almost All Organisms Have Some Form of Recombination
In molecular biology, the term recombination is used to refer to the physical cutting and joining of DNA molecules. We call this molecular recombination. This usage is quite distinct from recombination in the population genetic sense. On the one hand, new gene combinations can be produced by segregation, without any molecular recombination. On the other hand, new sequences can be produced by gene conversion and by recombination between repeated sequences (pp. 339–343). These involve molecular recombination within a single genome, and so from a population genetic point of view, we classify them as generating special kinds of mutations, rather than as mechanisms of genetic recombination.
Asexual Taxa Do Not Persist
Butlin (2002) reviews examples of ancient asexuals, including Darwinula (Fig. 23.8D).
Nunney (1989) discusses the role of species selection in maintaining sex.
Bell (1982, p. 328) discusses evidence for the hybrid origin of all vertebrate parthenogens.
Maynard Smith (1978) discusses asexual reproduction in dandelions (Taraxacum); see Hughes and Richards (1988) and Menken et al. (1995). Maynard Smith also contrasts arrhenotoky and thelytoky in insects.
Cullum (1997) compares fitness components in sexual and asexual Cnemidophorus and reviews earlier literature (Fig. 23.8A).
Vrijenhoek (1994) reviews unisexual fish, including Poeciliopsis (Fig. 23.8).
Parthenogenesis in Antennaria (Fig. 23.8C) is reviewed by Bierzychudek (1987).
Smith et al. (2006) give evidence for sexual reproduction in ostracods (Fig. 23.8D).
Welch and Meselson (2000) and Welch et al. (2004a,b) test the ancient asexuality of the bdelloid rotifers by comparing sequences at homologous loci (more information is provided ). In sexual populations, the two copies of each gene in a diploid trace back to a common ancestor approximately 2Ne generations ago and so differ by 4Neµ mutations on average (p. 426). However, in asexual organisms that produce genetically identical offspring, the two copies trace back much further, to a common ancestor that lived some time before asexuality was established. Welch et al. (2000, 2004a,b) found extremely high sequence divergence between homologous genes in bdelloid rotifers, consistent with . However, recent work suggests the possibility that this may be explained by tetraploidy instead.
Sex and Recombination Have Physiological, Genetic, and Evolutionary Costs
Several kinds of mutations are induced by meiosis: failure to repair the double-stranded breaks induced during molecular recombination; crossing over between nonhomologous sequences, which leads to insertions and deletions; and incorrect segregation of chromosomes. See pages 338–343.
Recombination load is reviewed by Charlesworth and Barton (1996); the only experimental data come from Mukai and Yamaguchi (1974) and Charlesworth and Charlesworth (1975).
When selection maintains stable linkage disequilibrium, modifiers that reduce recombination between favorable combinations of alleles will increase. This is known as the reduction principle (Feldman et al. 1996). Examples of such selection include the reduction in recombination responsible for evolution of Y chromosomes (p. 682), the tight linkage of components of segregation distorters (p. 590), and supergenes in Batesian mimics (p. 507).
Bernstein (1977) and Bernstein et al. (1985, 1988) have suggested that the function of diploidy, recombination, and sex is to repair double-stranded damage. Their argument is that diploidy is necessary so that an intact sequence is available to repair damage to the other strand and that crossing over is a side effect of repair. Thus far, the argument does not require sex or meiosis: Cells could maintain multiple reserve copies of the genome as insurance against damage without going to the trouble of mating. Bernstein et al.’s argument is that somatic crossing over causes sections of chromosome to become homozygous, which unmasks deleterious recessive alleles (Fig. WN23.7). Sex is therefore needed to restore heterozygosity, by bringing together gametes that carry different recessive alleles. Note, however, that although this argument may play a role in the origin of sex and recombination, it does not explain their current prevalence.
Sex Affects a Population Only If Alleles Associate Nonrandomly
Weissman’s (1889) argument is reviewed by Burt (2000).
We have discussed the effects of both segregation at a single locus and recombination between loci. However, the latter is likely to be more important for the evolution of sex, and most of the literature concentrates on recombination rather than segregation. We will follow the same course, but it is worth bearing in mind that many of the arguments we make about recombination and linkage disequilibrium carry over to segregation and deviations from Hardy–Weinberg proportions. See Kirkpatrick and Jenkins (1989).
Evans and Steinsaltz (2007) model the random segregation of mutational damage among offspring. They show that there is an optimal level of variation in damage, which maximizes the growth rate of the population. This argument is similar to Weissman’s argument that sex and recombination are favored because they generate variation that facilitates selection.
Competition Between Relatives Can Favor Genotypic Diversity
Evidence that genetic diversity leads to greater productivity comes from an experiment on Drosophila melanogaster larvae (Perez-Tome and Toro 1982). Two sets of vials were compared. In one set, eggs were laid by ten females that had all mated with the same male, whereas in the other set, ten females had mated with ten different males. Overall, survival was higher in the heterogeneous series. Fowler and Partridge (1986) criticized this experiment on the grounds that the effect could be due to variation in fertility. However, Martin et al. (1988) repeated the result, although only under the harshest conditions.
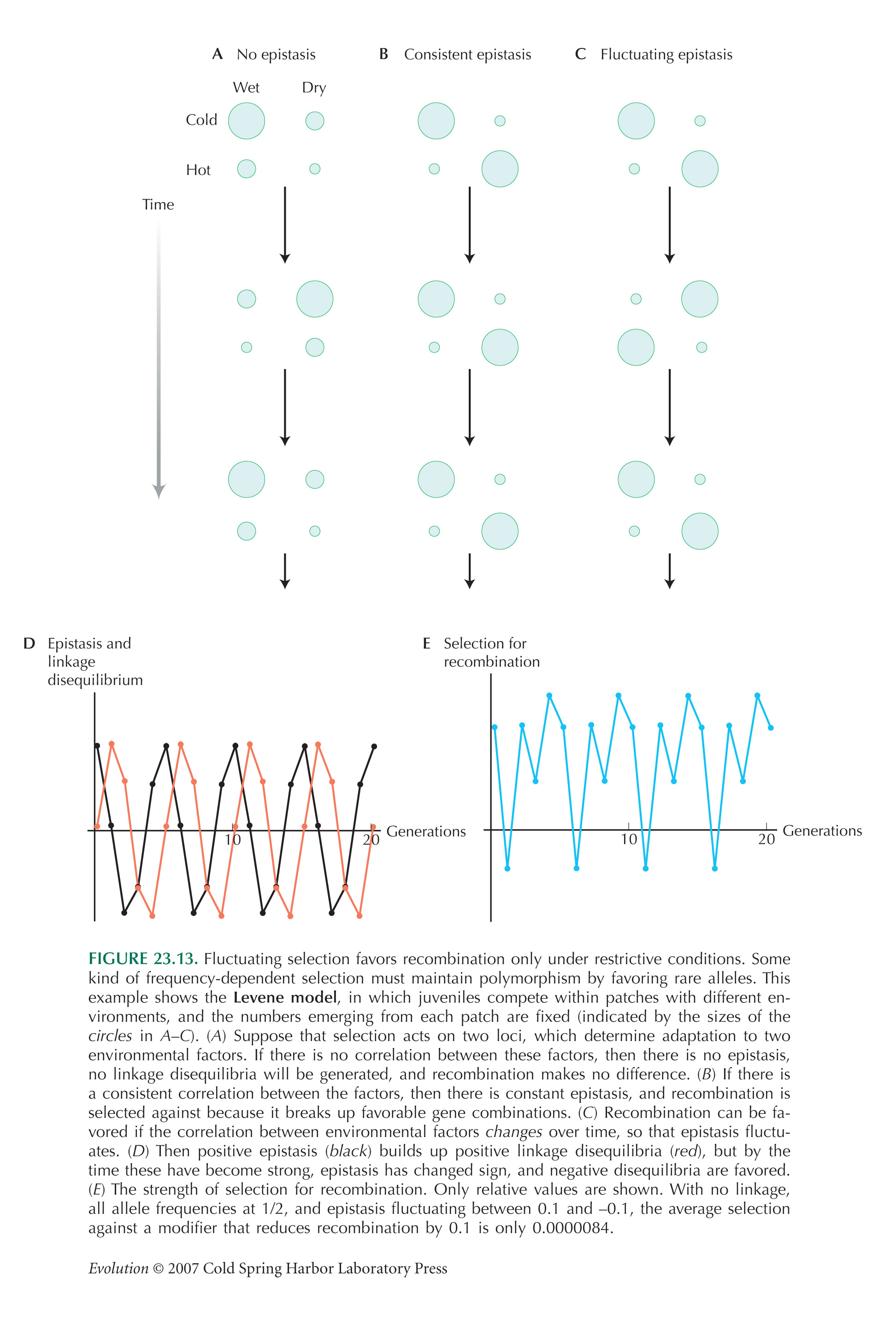
Fluctuating Environments Do Not Generally Favor Sex and Recombination
Figure 23.13D shows the selection for a modifier, from Equation 19 of Barton (1995); it is assumed that there is no directional selection, so that allele frequencies stay constant.
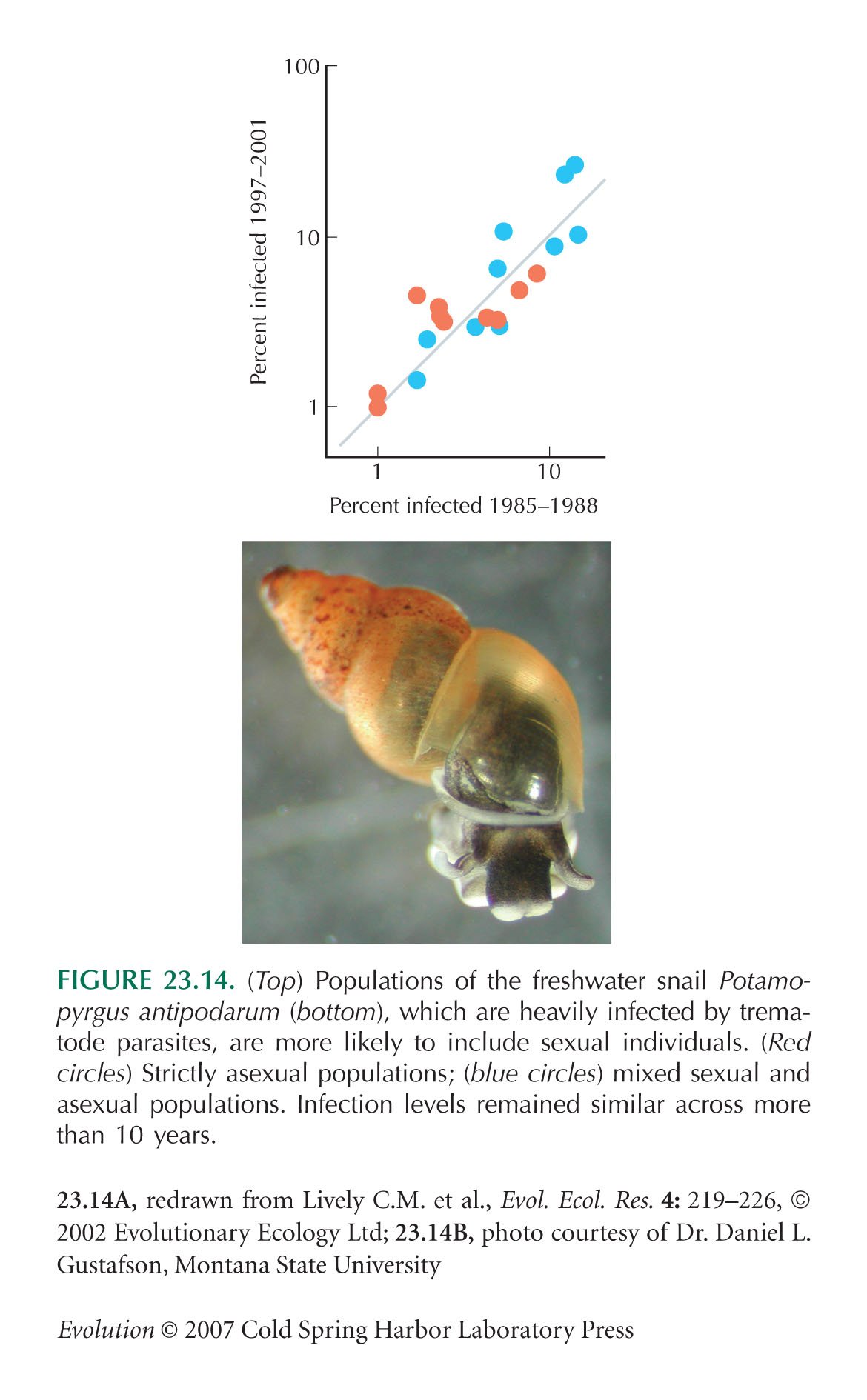
The Potamopyrgus example of Figure 23.14 is from Lively and Jokela (2002).
Recombination Increases the Additive Variance in Fitness When There Is Negative Linkage Disequilibrium
Whenever there is epistasis (reflected in a non-additive component of the variance in fitness), recombination causes an immediate reduction in fitness (i.e., a recombination load). See Barton (1995) and Barton and Charlesworth (1998).
Selection can also build up associations less directly. If the population is subdivided into local demes and if there is a negative correlation across demes between the strength of selection at different loci, then migration will generate negative associations and can drive the evolution of recombination. Again, though, it is hard to see why such negative correlations should exist and there is no evidence that they do. For further discussion of the role of migration in selection for recombination, see Otto and Lenormand (2002), Lenormand and Otto (2000), and Martin et al. (2005).
Negative Linkage Disequilibria Can Be Generated by Epistasis or by Drift
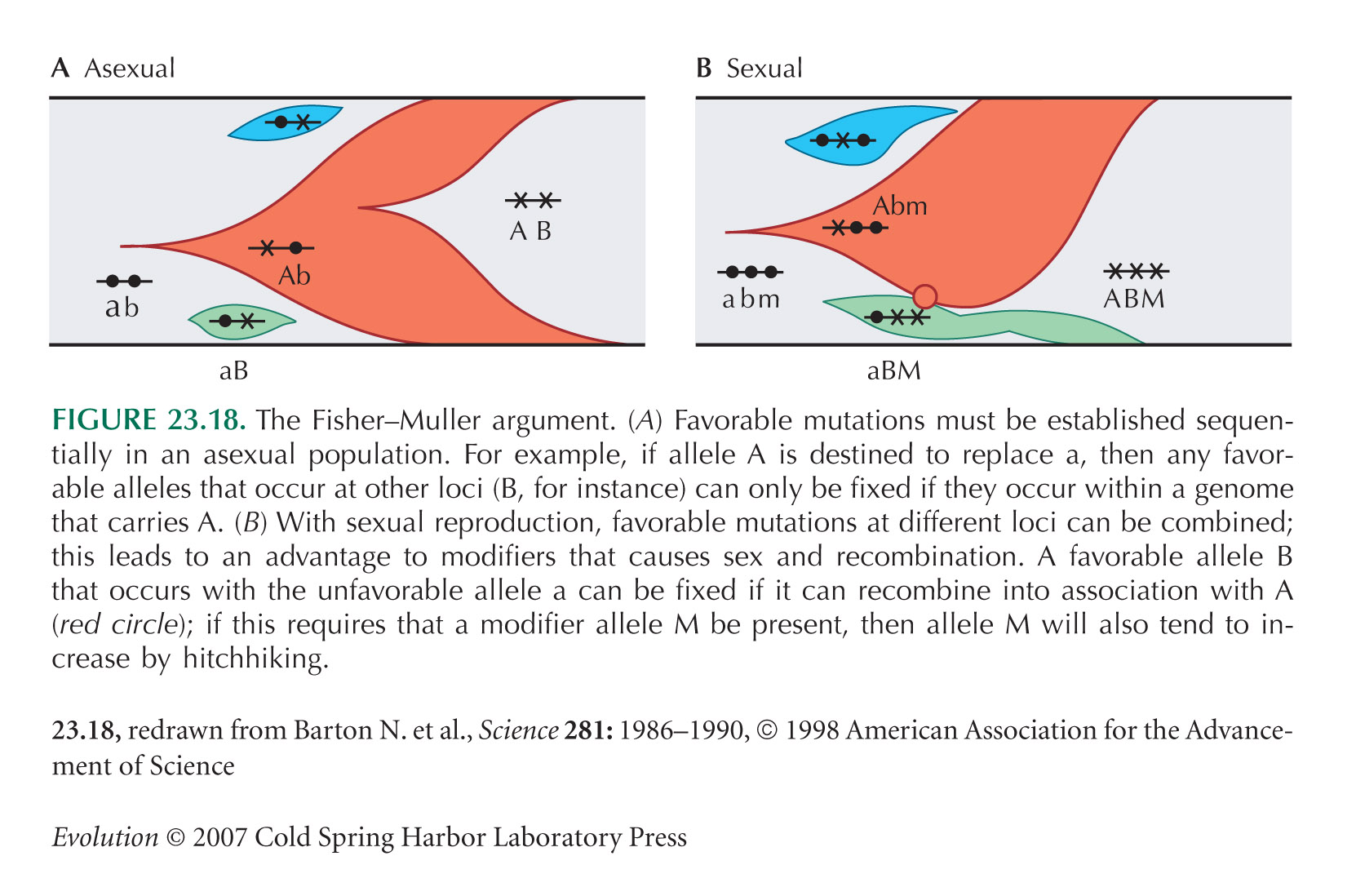
The diagram of the Fisher–Muller argument, Figure 23.18, is from Barton and Charlesworth (1998, Fig. 4), after Fisher (1930) and Muller (1932).
Interference between favorable mutations can be seen directly in bacterial populations. If a population is initially homogeneous, having grown from a single cell, then the frequency of some neutral allele will increase through mutation, µ, with a low frequency that increases linearly with time, t, as µt. When a favorable mutation sweeps through the population, it will almost certainly arise in the common genetic background and will sweep the marker out. Thus, the marker will show a sawtooth pattern, in which each selective sweep can be identified. This pattern is known as periodic selection. See Dykhuizen (1990).
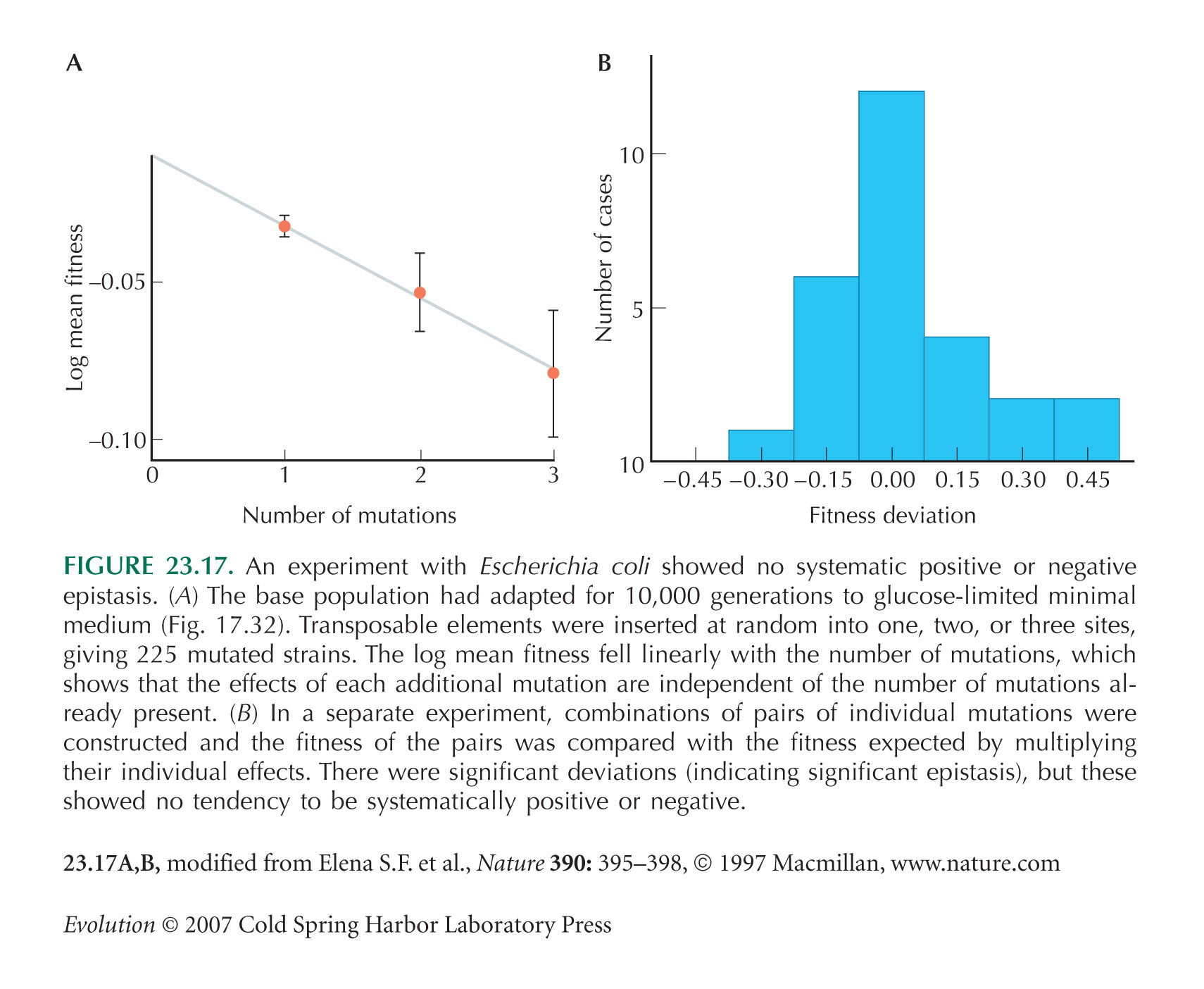
As noted by Elena and Lenski (1997), an additive scale in Figure 23.17A might be more appropriate. However, this makes virtually no difference to the results.
Random Linkage Disequilibria Amplify the Effects of Genetic Drift
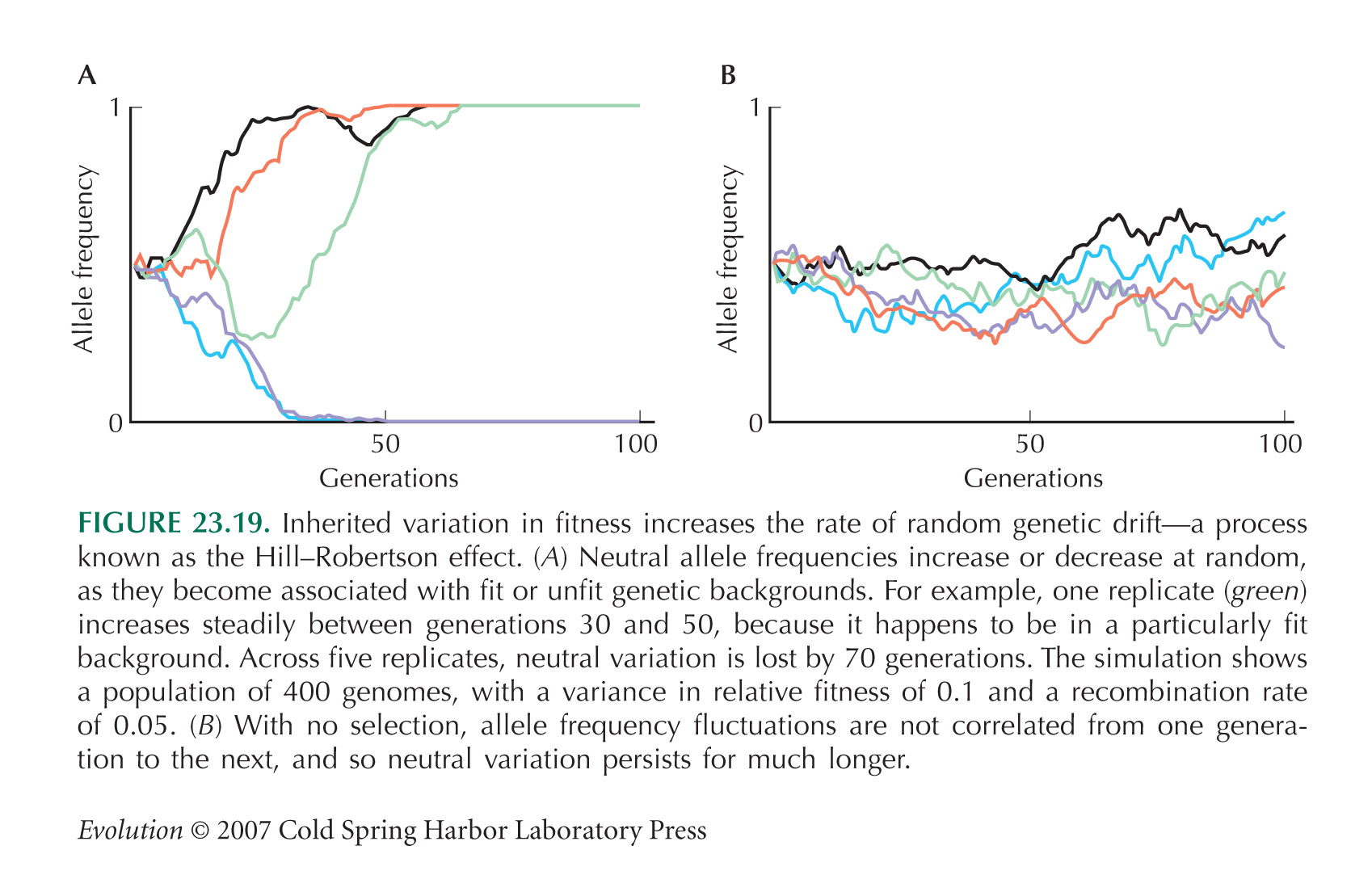
Hill and Robertson (1966) showed that selection increases the rate of random drift at linked loci and that this reduces the chances of fixation of favorable alleles.
Figure 23.19A shows a simulation in which the log fitness is mutated, by addition of a random Gaussian with standard deviation 0.1. In a population of 400 genomes, the variance in relative fitness settles to a long-term average of about 0.1. This model is a caricature of the true situation, in that all selection is concentrated at a single locus. For discussion of this kind of model, see Burt (2000).
Problem 23.9 shows how inherited variation in fitness, due to unlinked loci, causes four times the rate of random drift as does noninherited variation. This argument was made by Robertson (1966).
Favorable Alleles Are Brought Together by Recombination
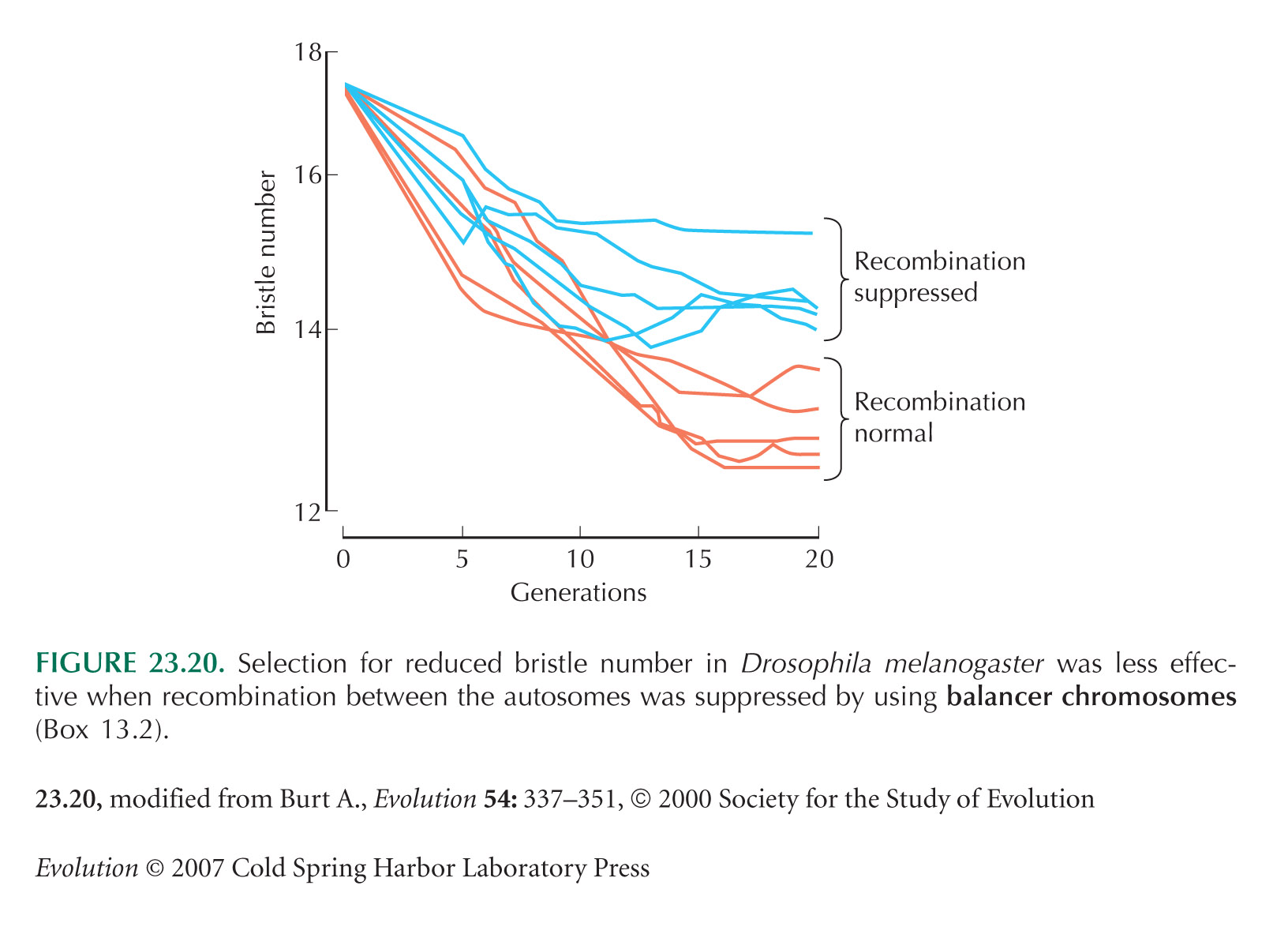
Figure 23.20 is based on an experiment by McPhee and Robertson (1970).
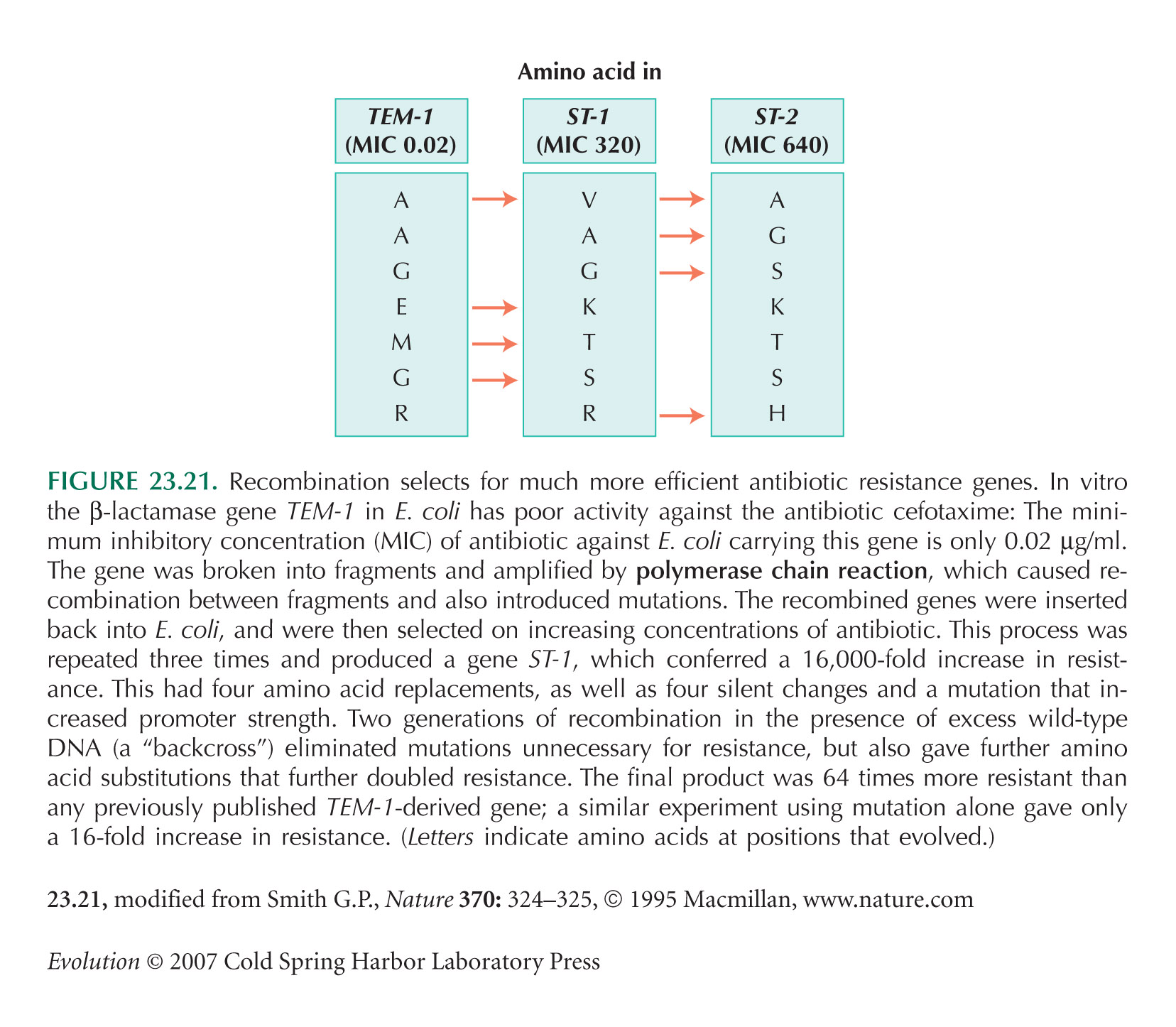
Figure 23.21 is based on an experiment by Stemmer (1994).
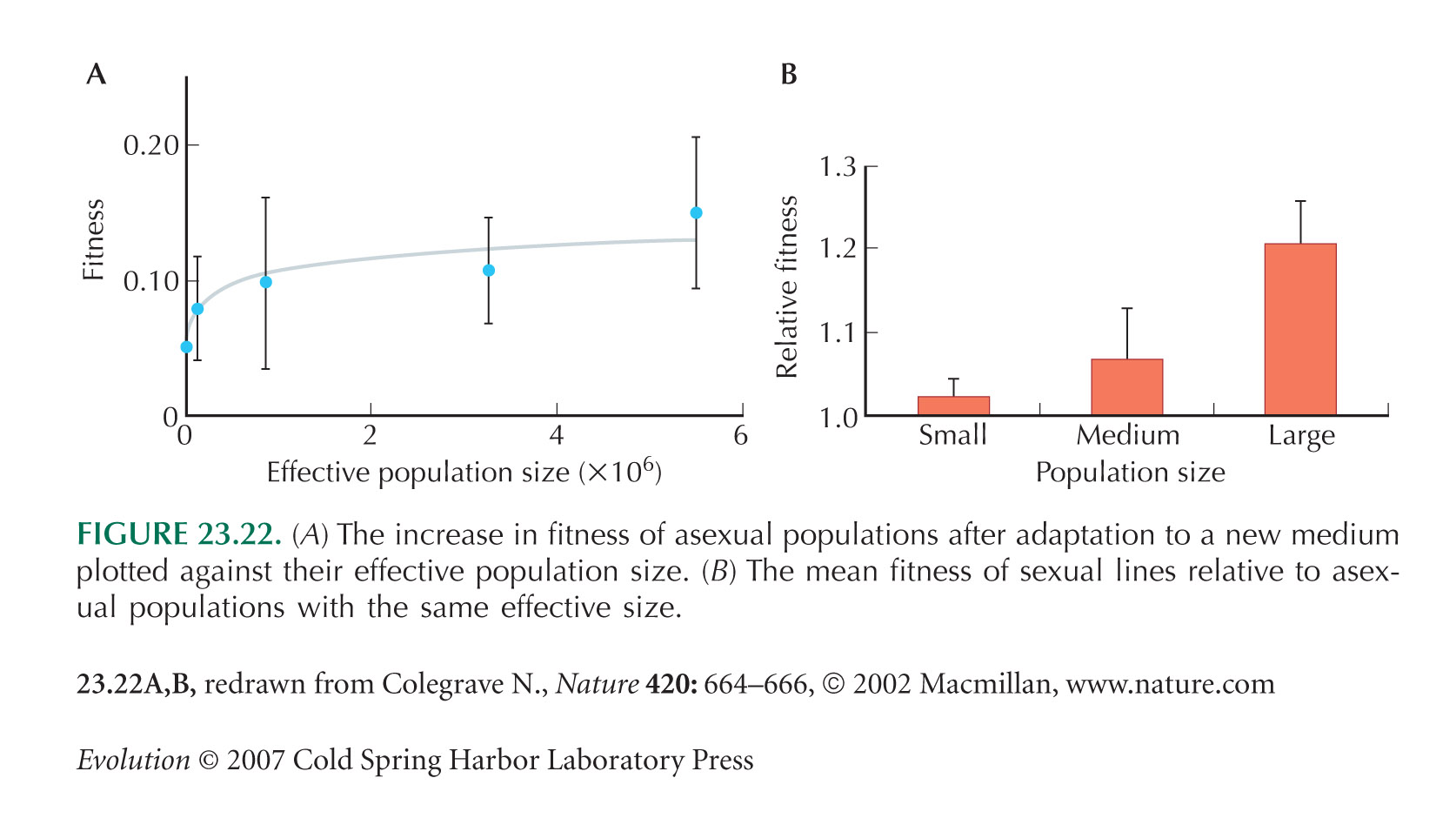
Figure 23.22 is from Colegrave (2002).
Poon and Chao (2003) designed an experiment that (like Fig. 23.22) explored the effect of population size on adaptation. However, adaptation was based on standing variation rather than on new mutations. We discuss their results .
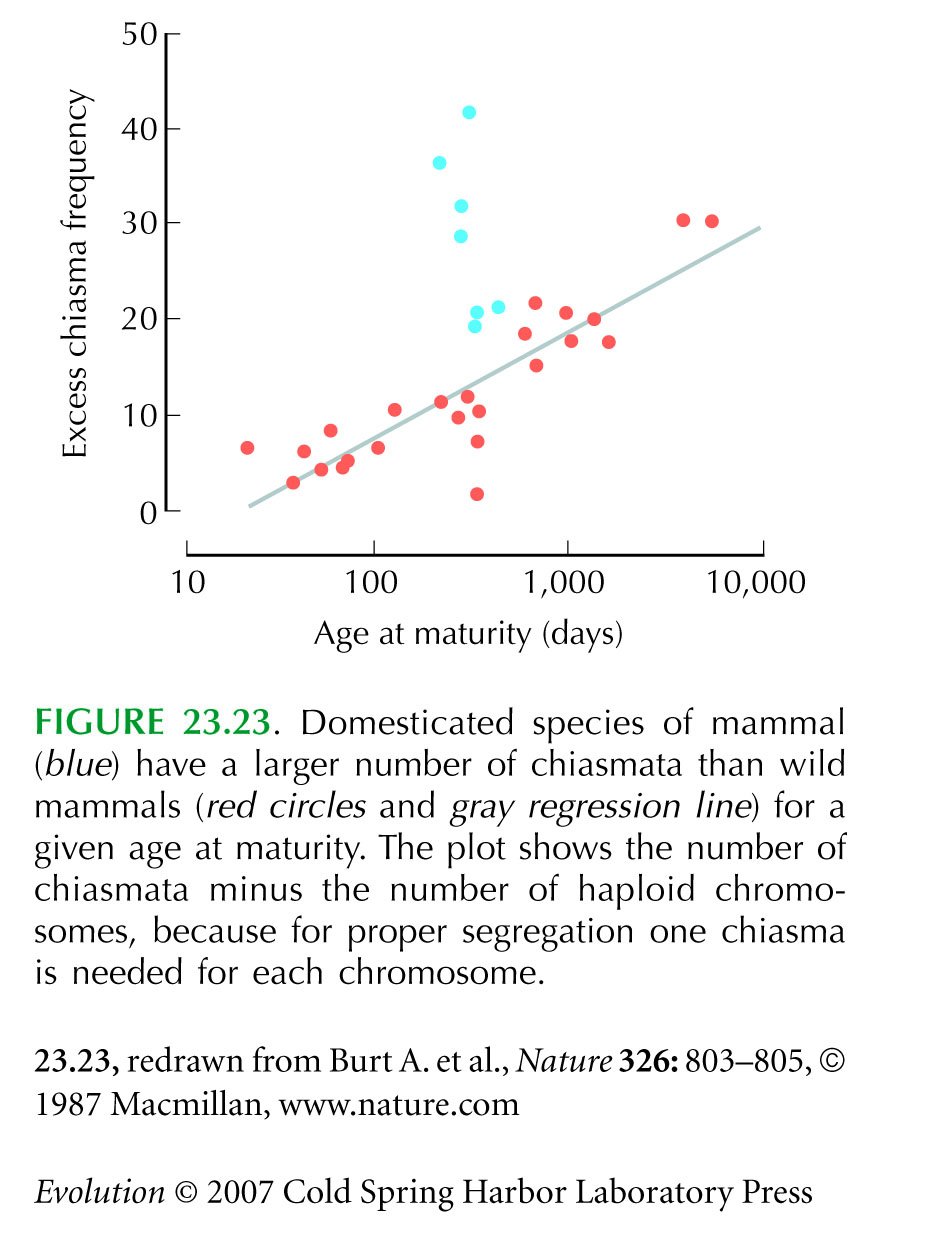
Figure 23.23 is from Burt and Bell (1987). See criticisms by Charlesworth et al. (1988).
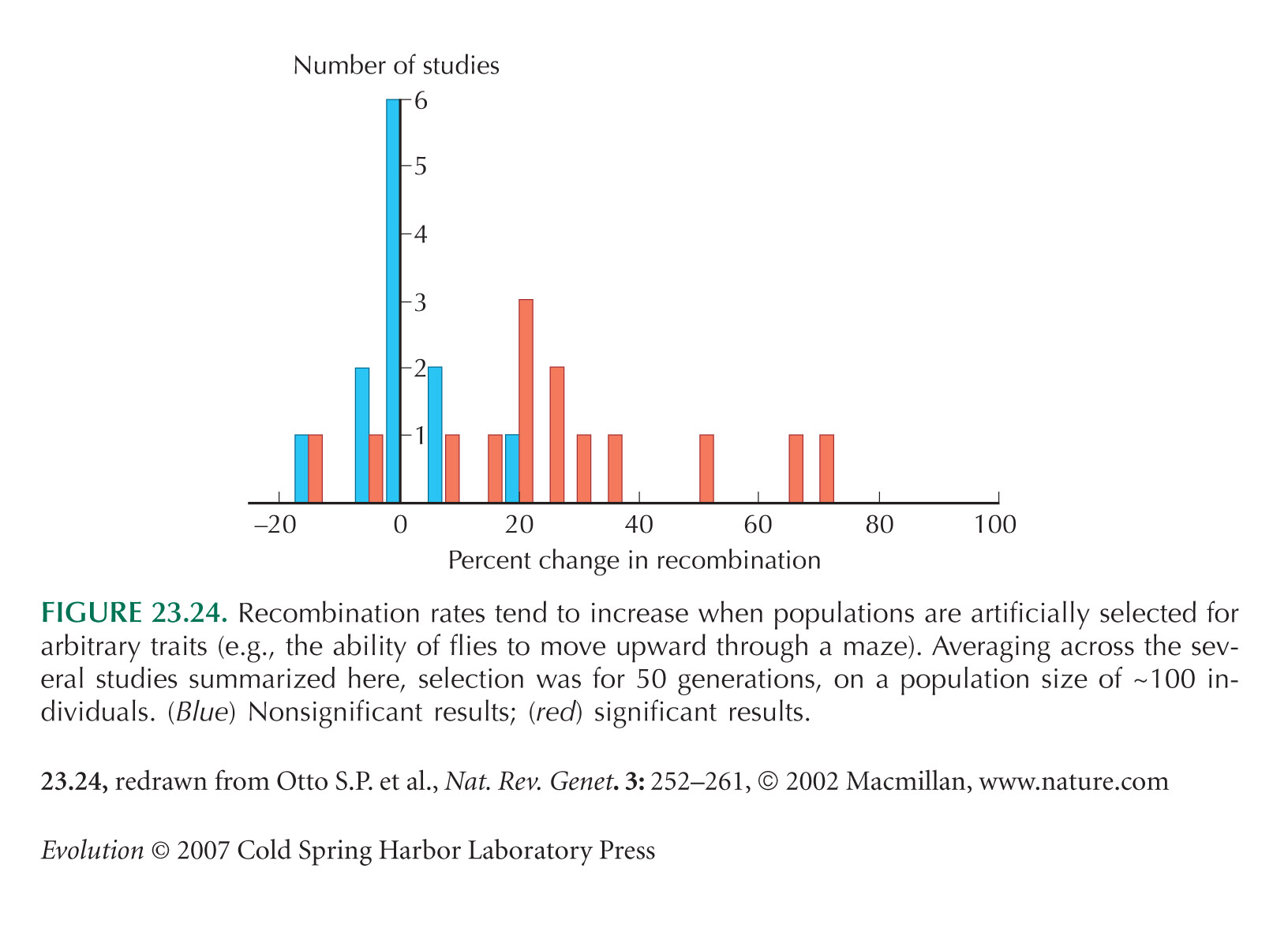
Figure 23.24 is from Otto and Lenormand (2002), based on data in Otto and Barton (2001).
In the Absence of Sex, Deleterious Alleles Accumulate
Kondrashov (1988) sets out the argument that if the total rate of deleterious mutation, U, is high, then the mutation load could only be tolerable with sexual reproduction and with negative epistasis. For discussion of the magnitude of U, see also Kondrashov (2000) and Haag-Liautard et al. (2007).
Muller (1964) described the random loss of the best genotype in an asexual population; Felsenstein (1974) termed this Muller’s ratchet and discussed its relation with the Hill–Robertson effect and the Fisher–Muller argument (Fig. 23.18). The process was analyzed mathematically by Haigh (1978). There is now a large literature on the ratchet: See, for example, Charlesworth and Charlesworth (1997) and Bachtrog and Gordo (2004).
Deleterious alleles may also accumulate as a result of positive selection. When a favorable mutation occurs, it will typically be in a genetic background carrying several (k ~ U/s) deleterious mutations. If the net fitness of this genome is less than that of the fittest class (i.e., S < ks, where S is the advantage of the new mutation), then the favorable mutation will be lost. However, if the selective advantage of the new mutation outweighs the disadvantage of the alleles with which it is associated, it can fix but will carry with it those deleterious alleles. Thus, selective sweeps cause the accumulation of deleterious alleles at other loci (Fig. WN23.9). See Peck (1994) and Johnson and Barton (2002).
Y Chromosomes Degenerate Because They Do Not Recombine
For reviews of the evolution of sex determination and sex chromosomes, see Charlesworth (1996, 2002). The evolution of dosage compensation is reviewed by Marin et al. (2000).
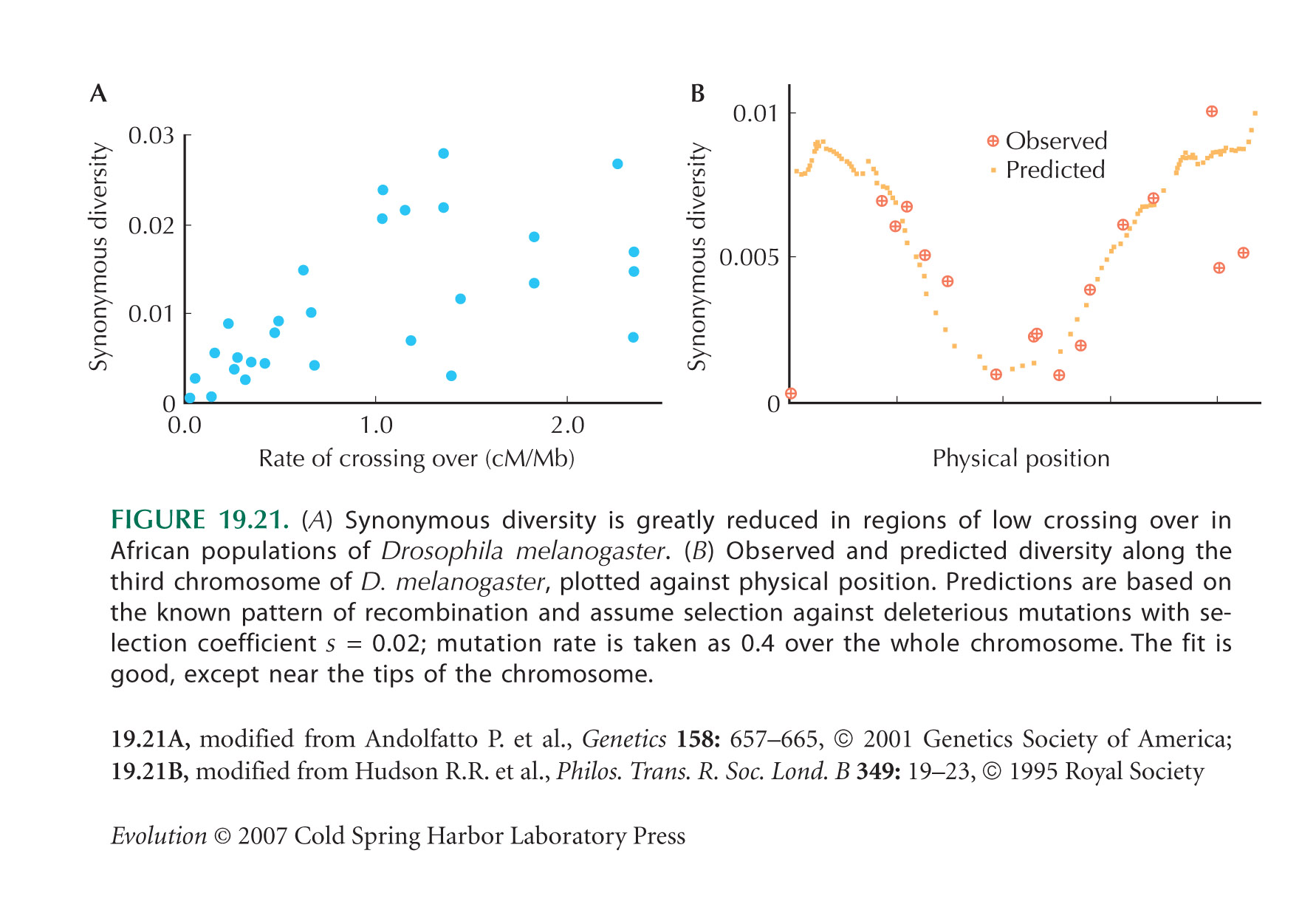
We saw in Figure 19.21 that regions of low recombination in D. melanogaster show reduced polymorphism, a pattern thought to be due to selection on linked loci. However, Payseur and Nachman (2000) and Spencer et al. (2006) find no correlation between recombination rate and diversity in the human genome. It may be that there is enough recombination in humans for interference from selected loci to be negligible; in this section, we are looking at the extreme case of zero recombination.
The accumulation of transposable elements near the centromeres may be due to the reduced efficiency of selection in regions of low recombination (p. 216).
Permanent heterozygosity means that recessive lethals can be fixed on the Y chromosome, because their function will be complemented by their homologs on the X chromosome. However, we have seen that recessive deleterious mutations are kept at low frequency primarily by their slight effects on heterozygous fitness (). Therefore, permanent heterozygosity is not a key factor in explaining the degeneration of Y chromosomes.
We give examples of Y-chromosome degeneration from Drosophila and humans .
The guppy example of Figure 23.27 is from Lahn et al. (2001, Fig. 4). See also Rice (1996).
Consequences of Sex
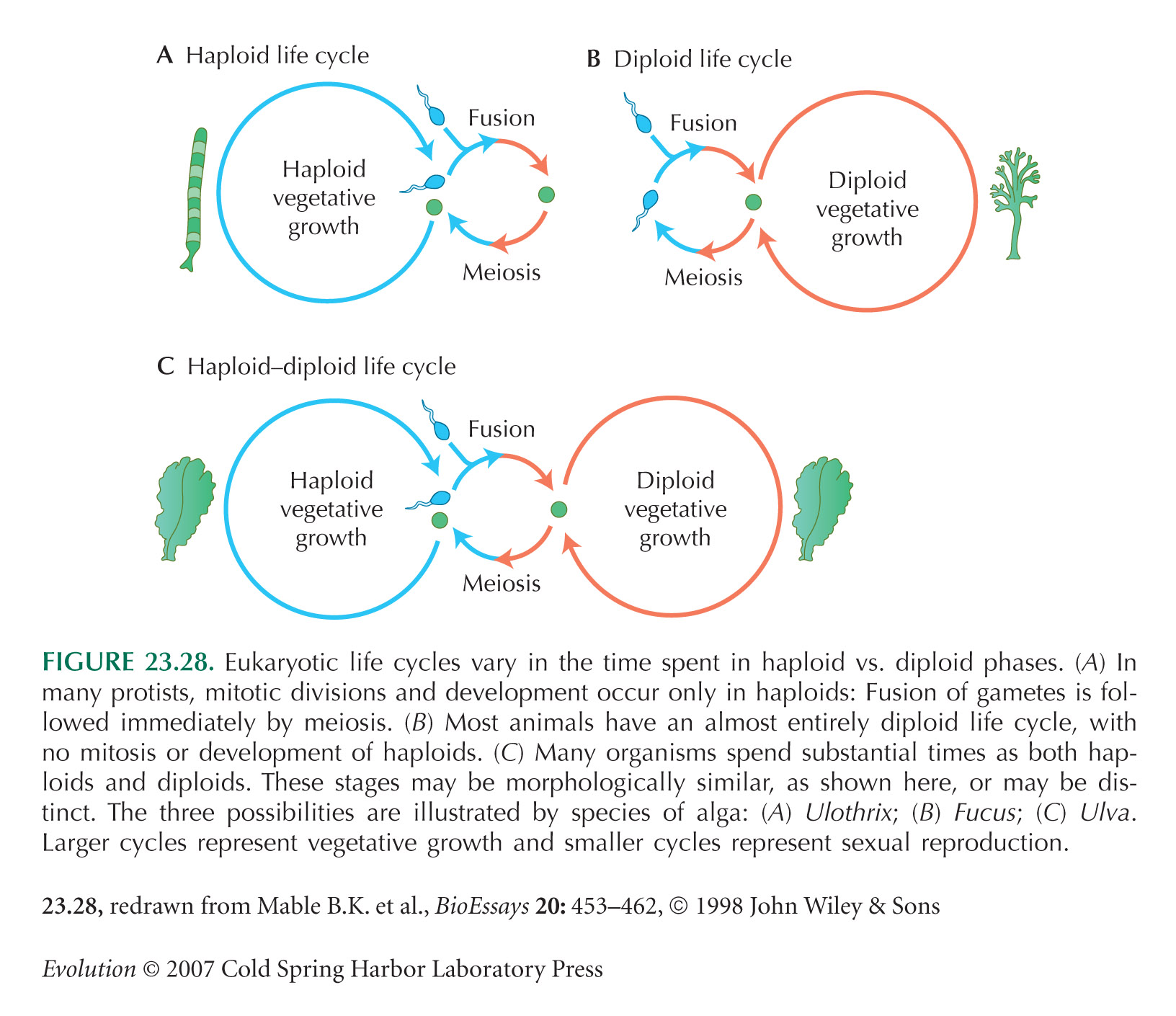
Diploid Life Cycles May Have Evolved to Mask Deleterious Alleles
The evolution of diploid life cycles is discussed by Maynard Smith and Szathmary (1995) and reviewed by Mable and Otto (1998).
The Pyrsonympha example is from Hollande and Carruette-Valentin (1970).
Where the haploid and diploid stages are distinct (Fig. 23.28C), they may specialize to exploit different ecological niches or one stage may be responsible for dispersal (Hughes and Otto 1999). The problem is similar to that of explaining why many insects have distinct larval and adult stages.
The Two Sexes Evolved through a Division of Labor between Gametes of Different Size
The evolution of anisogamy is discussed by Maynard Smith and Szathmary (1995) and Hoekstra (1987).
For the Volvocales comparison (Fig. 23.31), see Bell (1978), although the taxonomy is somewhat uncertain.
Many Mechanisms Have Evolved for Avoidance of Inbreeding
For reviews of plant mating systems, see Barrett (2002, 2003).
Stephenson and Winsor (1986) give an example of inbreeding avoidance through selective abortion. The idea that selective abortion is a means of eliminating selfed offspring was first suggested by Darwin (1862, 1883) as an explanation for the common excess of fruit production.
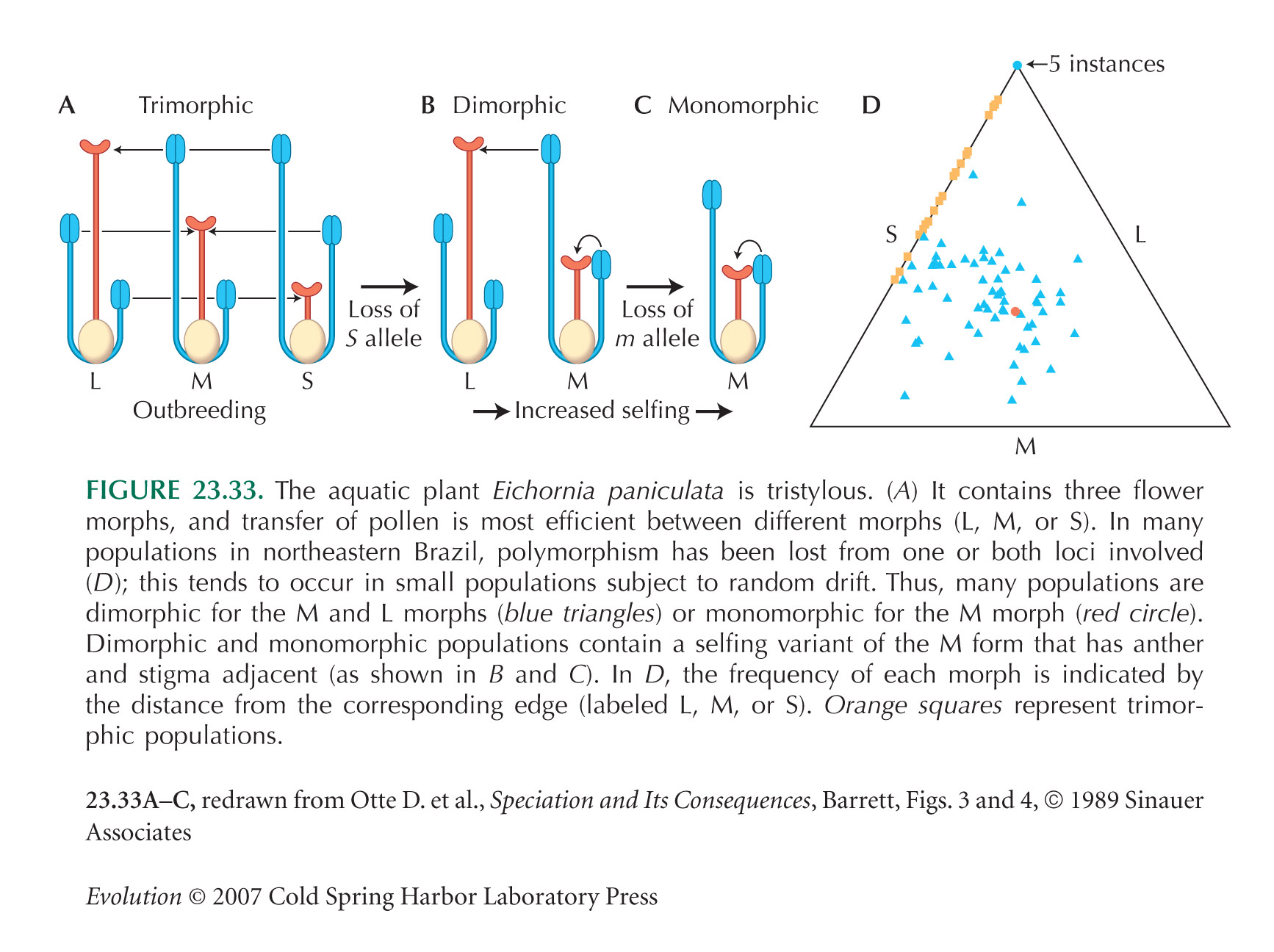
In Eichornia (Fig. 23.33), the pattern of loss of tristyly is determined by the genetic basis of the polymorphism. The S morph is produced by any plant that carries the S allele at one locus (S*/**, say), the M morph is produced by individuals carrying ss at one locus and the dominant M allele at the other (ssM*, say), and the L morph only by ssmm genotypes. Because frequency-dependent selection keeps these morphs at equal frequency, the dominant alleles tend to be rarer, and the recessive alleles more common, at equilibrium. Hence, the S morph tends to be lost first. Dimorphic populations then establish selfing variants of the M morph, which can go to fixation. See Barrett (1989).
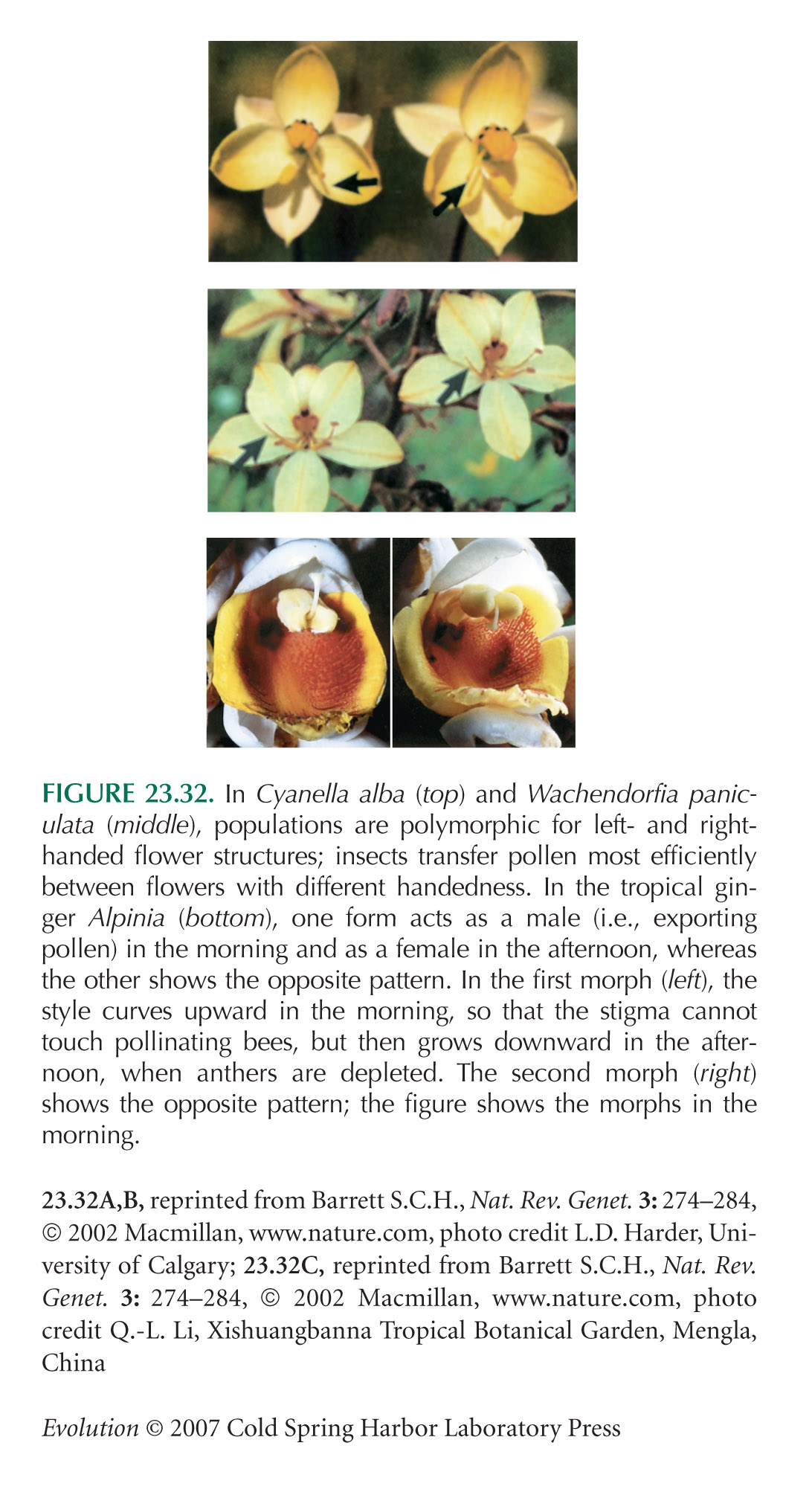
The examples of Figures 23.32 and 23.34 are from Barrett (2002).
The MHC (major histocompatibility complex) example is from Penn and Potts (1999).
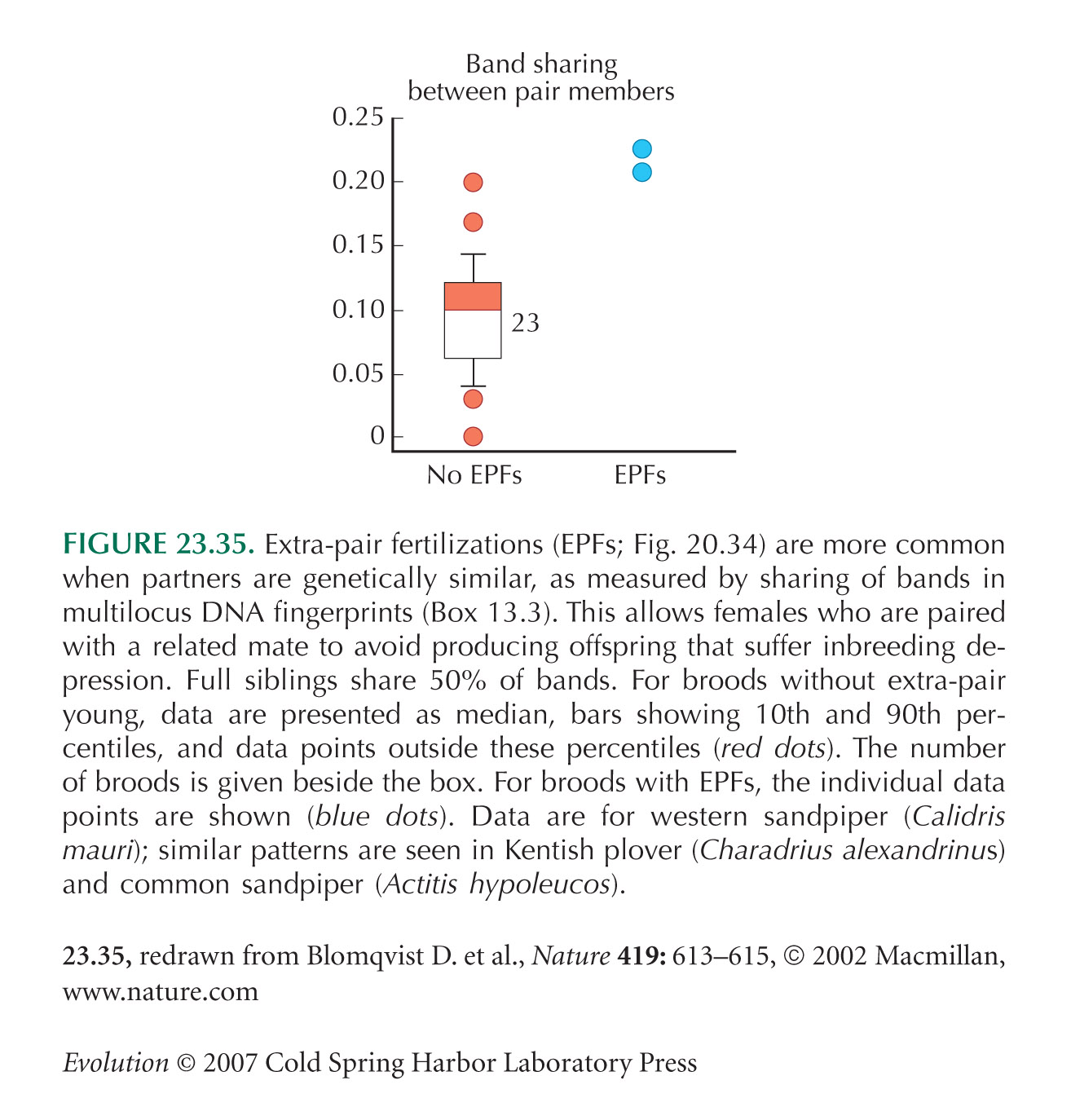
Extra-pair fertilization in birds (pp. 582–583) may be a means of avoiding inbreeding. Figure 23.35 is from Blomqvist et al. (2002, 2003).
Evolution of Evolvability
Dominance of Wild-type Alleles Is Only Weakly Selected
The evolution of dominance is reviewed by Mayo and Burger (1997). For historical background, see Provine (1986).
The Stentor example of Figure 23.37 is from Kirschner et al. (2000).
Charlesworth (1979) showed that in Drosophila, mutations of small effect are nearly additive, whereas those of large effect are recessive, which supports the physiological theory.
Orr (1992) shows that spontaneous mutations in Chlamydomonas reinhardtii are mostly recessive.
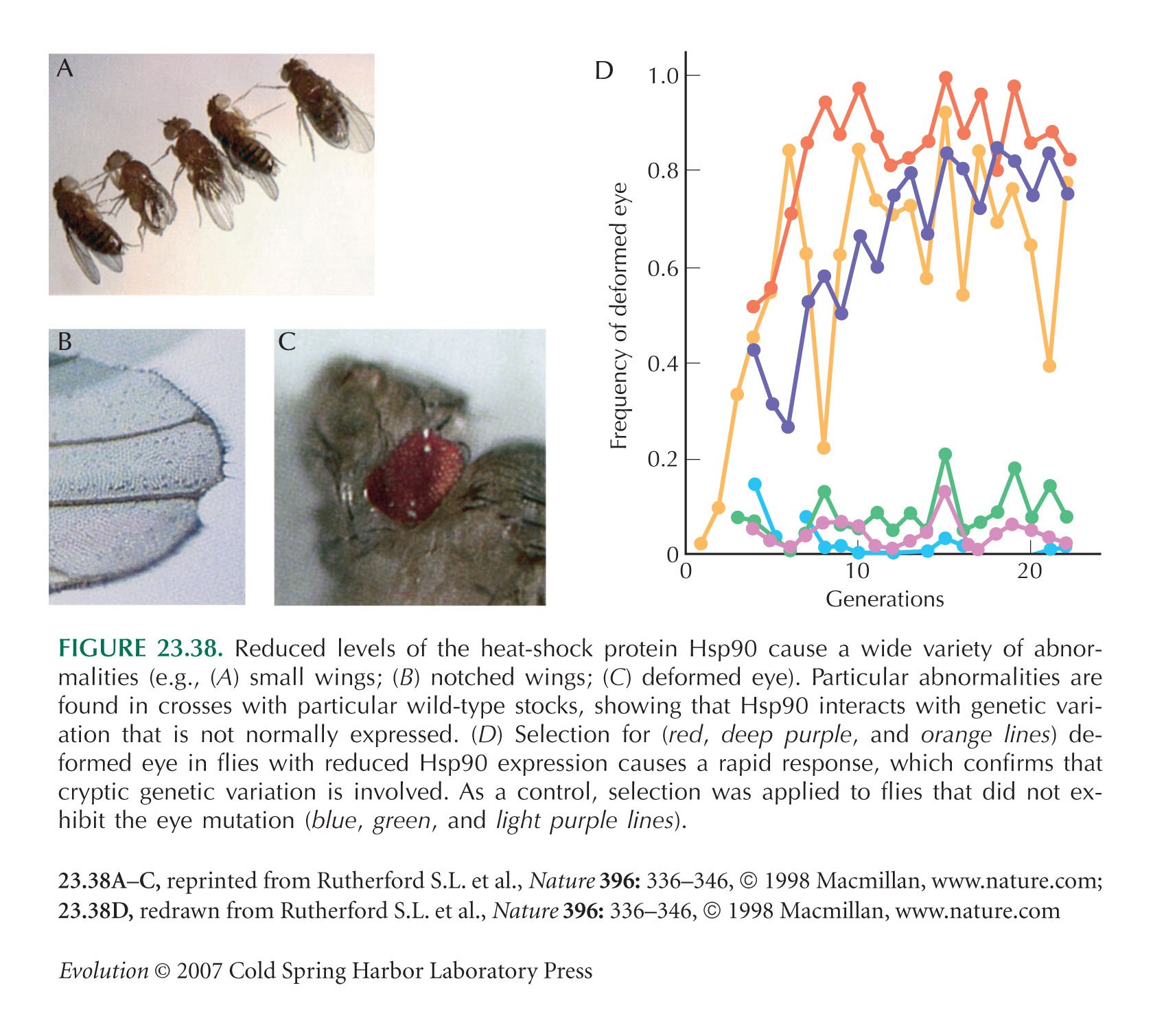
The Hsp90 example of Figure 23.38 is from Rutherford and Lindquist (1998); see also Queitsch et al. (2002). For discussion of the relation with Waddington’s experiments, see also Rutherford (2000).
Fares et al. (2002) show that GroEL masks deleterious mutations in E. coli. Their study was motivated by Moran’s (1996) suggestion that the high expression of GroEL in Buchnera compensates for deleterious mutations that have accumulated in this endosymbiont. GroEL seems not simply to give some general fitness advantage: GroEL overexpression had no deleterious effect on the ancestral E. coli when sufficient amino acids were present.
Robustness Can Evolve in Several Ways
There is a difficulty with strictly asexual populations, in that populations which evolve higher robustness will then accumulate a higher frequency of deleterious mutations and will in the long run gain no fitness advantage at all. (A similar problem arises with the evolution of diploid life cycles [pp. 683–685], which reduces the immediate mutation load by masking deleterious recessive mutations, but in the long run, doubles the load; Perrot et al. 1991, Kondrashov 1994.) Thus, evolution of robustness is easier to understand for sexual populations, where the increased mutation load is borne by the whole population and not just by those individuals that are more robust.
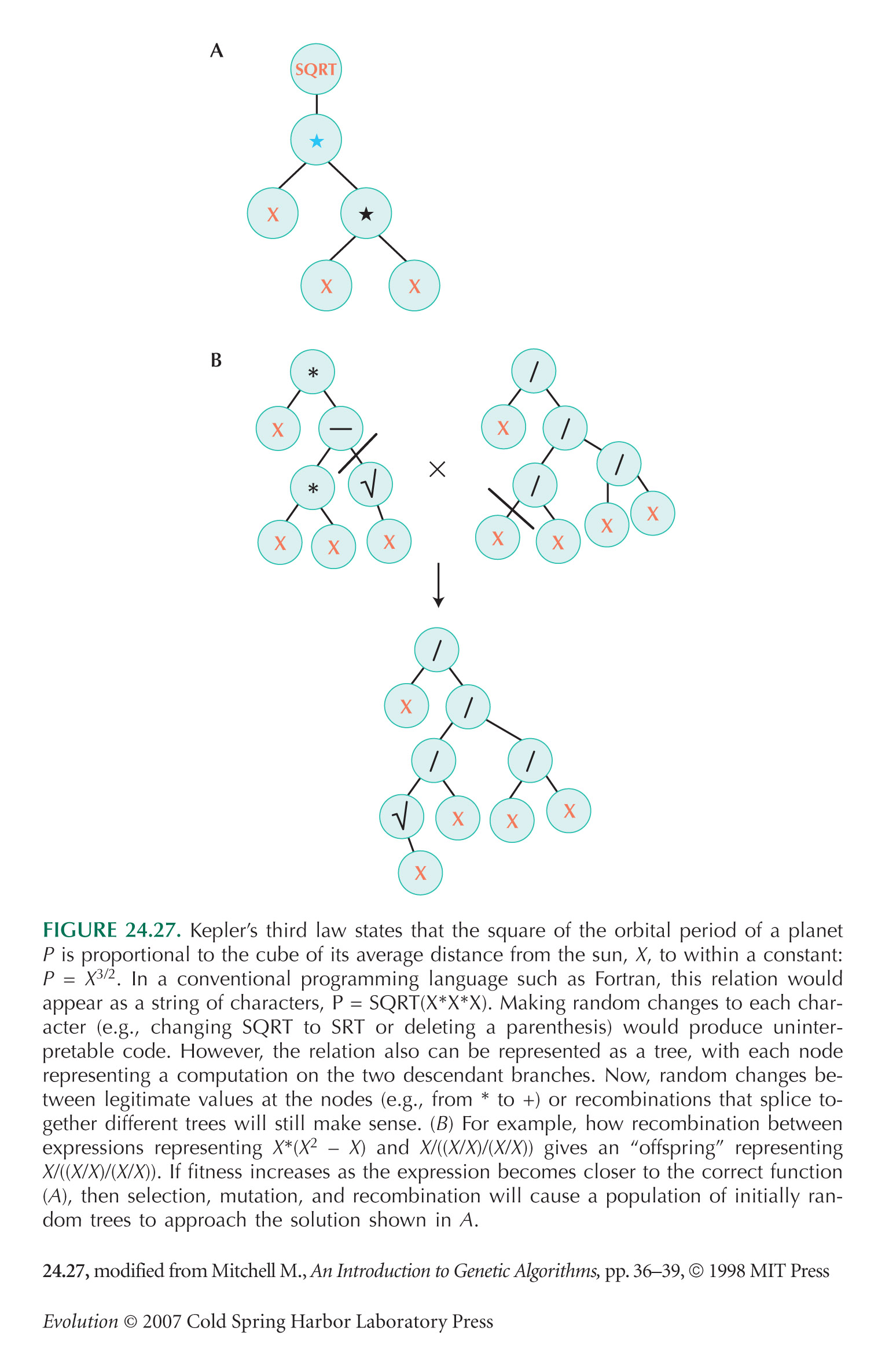
The evolvability of biological organisms is highlighted by the attempts of computer scientists to develop evolutionary algorithms through natural selection in silico (e.g., see Fig. 17.13). Success depends on recombination, but in most computer languages, recombining two working programs by cutting and pasting is disastrous. Evolutionary algorithms therefore require specially designed languages, which function even if disrupted by recombination (e.g., see Fig. 24.27).
References
Bachtrog D. 2003. Adaptation shapes patterns of genome evolution on sexual and asexual chromosomes in Drosophila. Nat. Genet. 34: 215–219.
Bachtrog D. and Gordo I. 2004. Adaptive evolution of asexual populations under Muller’s ratchet. Evolution 58: 1403–1413.
Ballard J.W. and Whitlock M. 2004. The incomplete natural history of mitochondria. Mol. Ecol. 13: 729–744.
Barrett S.C.H. 1989. Mating system evolution and speciation in heterostylous plants. In Speciation and its consequences (ed. D. Otte and A. Endler), pp. 257–283. Sinauer Press, Sunderland, Massachusetts.
Barrett S.C.H. 2002. The evolution of plant sexual diversity. Nat. Rev. Genet. 3: 274–284.
Barrett S.C.H. 2003. Mating strategies in flowering plants: The outcrossing-selfing paradigm and beyond. Philos. Trans. R. Soc. Lond. B Biol. Sci. 358: 991–1004.
Barton N.H. 1995. A general model for the evolution of recombination. Genet. Res. 65: 123–144.
Barton N.H. and Charlesworth B. 1998. Why sex and recombination? Science 281: 1986–1990.
Bell G. 1978. The evolution of anisogamy. J. Theor. Biol. 73: 247–270.
Bell G. 1982. The masterpiece of nature: The evolution and genetics of sexuality. University of California Press, Berkeley.
Bernstein H. 1977. Germline recombination may be primarily a manifestation of DNA repair processes. J. Theor. Biol. 69: 371–380.
Bernstein H., Byerly H.C., Hopf F.A., and Michod R. 1985. Genetic damage, repair, and the evolution of sex. Science 229: 1277–1281.
Bernstein H., Hopf F.A., and Michod R.E. 1988. Is meiotic recombination an adaptation for repairing DNA, producing genetic variation, or both? In The evolution of sex (ed. R.E. Michod and B.R. Levin), pp. 139–160. Sinauer Press, Sunderland, Massachusetts.
Bierzychudek P. 1987. Patterns in plant parthenogenesis. In The evolution of sex and its consequences (ed. S.C. Stearns), pp. 197–218. Birkhauser Verlag, Basel.
Blomqvist D., Andersson M., Kupper C., Cuthill I.C., Kis J., et al. 2002. Genetic similarity between mates and extra-pair parentage in three species of shorebirds. Nature 419: 613–615.
Blomqvist D., Andersson M., Kupper C., Cuthill I.C., Kis J., et al. 2003. Why do birds engage in extra-pair copulation? Reply. Nature 422: 833–834.
Brooks L.D. 1988. The evolution of recombination rates. In The evolution of sex (ed. R.E. Michod and B.R. Levin), pp. 87–105. Sinauer Press, Sunderland, Massachusetts.
Burt A. 2000. Sex, recombination and the efficacy of selection—Was Weissman right? Evolution 54: 337–351.
Burt A. and Bell G. 1987. Mammalian chiasma frequencies as a test of two theories of recombination. Nature 326: 803–805 and 330: 118.
Butlin R. 2002. The costs and benefits of sex: New insights from old asexual lineages. Nat. Rev. Genet. 3: 311–317.
Charlesworth B. 1979. Evidence against Fisher’s theory of dominance. Nature 278: 848–849.
Charlesworth B. 1996. The evolution of chromosomal sex determination and dosage compensation. Curr. Biol. 6: 149–162.
Charlesworth B. 2002. The evolution of chromosomal sex determination. In The genetics and biology of sex determination (ed. D. Chadwick and J. Goode), Novartis Foundation Symposium 244: 207–219; discussion 220–224, 253–257.
Charlesworth B. and Barton N.H. 1996. Recombination load associated with selection for increased recombination. Genet. Res. 77: 27–41.
Charlesworth B. and Charlesworth D. 1975. An experiment on recombination load in Drosophila melanogaster. Genet. Res. (Camb.) 25: 267–274.
Charlesworth B. and Charlesworth D. 1997. Rapid fixation of deleterious alleles can be caused by Muller’s ratchet. Genet. Res. 70: 63–74.
Charlesworth B. and Charlesworth D. 2000. The degeneration of Y chromosomes. Philos. Trans. R. Soc. Lond. B 355: 1563–1572.
Charlesworth D., Charlesworth B., and Marais G. 2005. Steps in the evolution of heteromorphic sex chromosomes. Heredity 95: 118–128.
Charlesworth B., Greenwood J., and Koella J.C. 1988. Comment on Burt and Bell, 1987. Nature 330: 118.
Colegrave N. 2002. Sex releases the speed limit on evolution. Nature 420: 664–666.
Coop G., Wen X., Ober C., Pritchard J.K., and Przeworski M. 2008. High-resolution mapping of crossovers reveals extensive variation in fine-scale recombination patterns among humans. Science 319: 1395–1398.
Cullum A.J. 1997. Comparisons of physiological performance in sexual and asexual whiptail lizards (genus Cnemidophorus): Implications for the role of heterozygosity. Am. Nat. 150: 24–47.
Darwin C. 1862. On the two forms, or dimorphic condition, in the species Primula, and on their remarkable sexual relations. J. Proc. Linn. Soc. Lond (Bot.) 6 (Read 21 November 1861): 77–96.
Darwin C. 1883. The variation of animals and plants under domestication, volumes I and II (2nd edition, revised). D. Appleton & Co., New York.
Dawson K.J. 1998. Evolutionarily stable mutation rates. J. Theor. Biol. 194: 143–158.
Drake J.W. and Holland J.J. 1999. Mutation rates among RNA viruses. Proc. Natl. Acad. Sci. 96: 13910–13913.
Drake J.W., Charlesworth B., Charlesworth D., and Crow J.F. 1998. Rates of spontaneous mutation. Genetics 148: 1667–1686.
Dykhuizen D.E. 1990. Experimental studies of natural selection in bacteria. Annu. Rev. Ecol. Syst. 21: 373–398.
Elena S.F. and Lenski R.E. 1997. Test of synergistic interactions among deleterious mutations in bacteria. Nature 390: 395–298.
Evans S.N. and Steinsaltz D. 2007. Damage segregation at fissioning may increase growth rates: A superprocess model. Theor. Popul. Biol. 71: 473–490.
Eyre-Walker A., Smith N.H., and Maynard Smith J. 1999. How clonal are human mitochondria? Proc. R. Soc. Lond. B 266: 477–483; 2041–2042.
Fares M.A., Ruiz-Gonzalez M.X., Moya A., Elena S.F., and Barrio E. 2002. GroEL buffers against deleterious mutations. Nature 417: 398.
Feldman M.W., Otto S.P., and Christiansen F.B. 1996. Population genetic perspectives on the evolution of recombination. Annu. Rev. Genet. 30: 261–295.
Felsenstein J. 1974. The evolutionary advantage of recombination. Genetics 78: 737–756.
Fisher R.A. 1930. The genetical theory of natural selection. Oxford University Press, Oxford.
Fowler K. and Partridge L. 1986. Variation in male fertility explains an apparent effect of genotype diversity on success in larval, competition in Drosophila melanogaster. Heredity 57: 31–36.
Giraud A., Matic I., Tenaillon O., Clara A., Radman M., et al. 2001. Costs and benefits of high mutation rates: Adaptive evolution of bacteria in the mouse gut. Science 291: 2606–2608.
Gladyshev E. and Meselson M.S. 2008. Extreme resistance of bdelloid rotifers to ionizing radiation. Proc. Natl. Acad. Sci. 105: 5139–5144.
Haag-Liautard C., Dorris M., Maside X., Macaskill S., Halligan D.L., Charlesworth B., and Keightley P.D. 2007. Direct estimation of per nucleotide and genomic deleterious mutation rates in Drosophila. Nature 445: 82–85.
Haigh J. 1978. The accumulation of deleterious genes in a population—Muller’s ratchet. Theor. Popul. Biol. 14: 251–267.
High N.J., Deadman M.E., and Moxon E.R. 1993. The role of a repetitive DNA motif (5′-CAAT-3′) in the variable expression of the Haemophilus influenzae lipopolysaccharide epitope αGal(1-4)βGal. Mol. Microbiol. 9: 1275–1282.
Hill W.G. and Robertson A. 1966. The effect of linkage on limits to artificial selection. Genet. Res. 8: 269–294.
Hoekstra R.F. 1987. The evolution of sexes. In The evolution of sex and its consequences (ed. S.C. Stearns), pp. 59–92. Birkhauser Verlag, Basel.
Hollande A. and Carruette-Valentin J. 1970. La lignée des Pyrsonymphines et les caractères infrastructuraux communs aux genres Opisthomitus, Oxymonas, Saccinobacculus, Pyrsonympha et Streblomastix. CR Acad. Sci. Paris 270: 1587–1590.
Hughes J.S. and Otto S. P. 1999. Ecology and the evolution of biphasic life cycles. Am. Nat. 154: 306–320.
Hughes J. and Richards A.J. 1988. The genetic structure of populations of sexual and asexual Taraxacum (dandelions). Heredity 60: 161–172.
Innan H. and Nordborg M. 2002. Recombination or mutational hot spots in human mtDNA? Mol. Biol. Evol. 19: 1122–1127.
Johnson T. and Barton N.H. 2002. The effect of deleterious alleles on adaptation in asexual populations. Genetics 162: 395–411.
Kirkpatrick M. and Jenkins C.D. 1989. Genetic segregation and the maintenance of sexual reproduction. Nature 339: 300–301.
Kirschner M., Gerhart J., and Mitchison T. 2000. Molecular “vitalism.” Cell 100: 79–88.
Kondrashov A.S. 1988. Deleterious mutations and the evolution of sexual reproduction. Nature 336: 435–441.
Kondrashov A.S. 1994. The asexual ploidy cycle and the origin of sex. Nature 370: 213–216.
Kondrashov A.S. 2000. Sex and U. Trends Genet. 17: 75–77.
Kroll J.S., Loynds B.M., and Moxon E.R. 1991. The Haemophilus influenzae capsulation gene cluster: A compound transposon. Mol. Microbiol. 5: 1549–1560.
Ladoukakis E.D. and Zouros E. 2001. Direct evidence for homologous recombination in mussel (Mytilus galloprovincialis) mitochondrial DNA. Mol. Biol. Evol. 18: 1168–1175.
Lahn B.T. and Page B.T. 1999. Four evolutionary strata on the human Y chromosome. Science 286: 964–967.
Lahn B.T., Pearson N.M., and Jegalian K. 2001. The human Y chromosome, in the light of evolution. Nat. Rev. Genet. 2: 207–216.
Lenormand T. and Otto S.P. 2000. The evolution of recombination in a heterogeneous environment. Genetics 156: 423–38.
Levinson G. and Gutman G.A. 1987. Slipped-strand mispairing: A major mechanism for DNA sequence evolution. Mol. Biol. Evol. 4: 203–221.
Lively C.M. and Jokela J. 2002. Temporal and spatial distributions of parasites and sex in a freshwater snail. Evol. Ecol. Res. 4: 219–226.
Lynch M. 1997. Mutation accumulation in nuclear, organelle, and prokaryotic transfer RNA genes. Mol. Biol. Evol. 14: 914–925.
Lynch M. and Blanchard J.L. 1998. Deleterious mutation accumulation in organelle genomes. Genetica 103: 29–39.
Mable B.K. and Otto S.P. 1998. The evolution of life cycles with haploid and diploid phases. BioEssays 20: 453–462.
Macaulay V., Richards M., and Sykes B. 1999. Mitochondrial DNA recombination—No need to panic. Proc. R. Soc. Lond. B 266: 2037–2039.
Marin I., Siegal M.L., and Baker B.S. 2000. The evolution of dosage compensation mechanisms. BioEssays 22: 1106–1114.
Martin G., Otto S.P., and Lenormand T. 2005. Selection for recombination in structured populations. Genetics 172: 593–609.
Martin M.J., Perez-Tome J.M., and Toro M.A. 1988. Competition and genotypic variability in Drosophila melanogaster. Heredity 69: 119–124.
Matic I., Radman M., Taddei F., Picard B., Doit C., et al. 1997. Highly variable mutation rates in commensal and pathogenic Escherichia coli. Science 277: 1833–1834.
Maynard Smith J. 1978. The evolution of sex. Cambridge University Press, Cambridge.
Maynard Smith J. and Szathmáry E. 1995. The major transitions in evolution. W.H. Freeman, Oxford.
Mayo O. and Burger R. 1997. The evolution of dominance: a theory whose time has passed? Biol. Rev. 72: 97–110.
McAllister B.F. and Charlesworth B. 1999. Reduced sequence variability on the neo-Y chromosome of Drosophila americana americana. Genetics 153: 221–233.
McPhee H.C. and Robertson A. 1970. The effect of suppressing crossing-over on the response to selection in D. melanogaster. Genet. Res. 16: 1–16.
Menken S.B.J., Smit E., and Den Nuis J.C.M. 1995. Genetical population structure in plants: Gene flow betwen diploid sexual and triploid asexual dandelions (Taraxacum section Ruderalia). Evolution 49: 1108–1118.
Moran N.A. 1996. Accelerated evolution and Muller’s rachet in endosymbiotic bacteria. Proc. Natl. Acad. Sci. 93: 2873–2878.
Moxon E.R., Rainey P.B., Nowak M.A., and Lenski R.E. 1994. Adaptive evolution of highly mutable loci in pathogenic bacteria. Curr. Biol. 4: 24–33.
Muller H.J. 1932. Some genetic aspects of sex. Am. Nat. 66: 118–138.
Muller H.J. 1964. The relation of recombination to mutational advance. Mutat. Res. 1: 2–9.
Mukai T. and Yamaguchi O. 1974. The genetic structure of natural populations of D. melanogaster. XI Genetic variability in a local population. Genetics 76: 339–366.
Nothel H. 1987. Adaptation of Drosophila melanogaster populations to high mutation pressure: Evolutionary adjustment of mutation rates. Proc. Natl. Acad. Sci. 84: 1045–1049.
Nunney L. 1989. The maintenance of sex by group selection. Evolution 43: 245–257.
Orr H.A. 1992. A test of Fisher’s theory of dominance. Proc. Natl. Acad. Sci. 88: 1413–1415.
Otto S.P. and Barton N.H. 2001. Selection for recombination in small populations. Evolution 55: 1921–1931.
Otto S. and Lenormand T. 2002. Resolving the paradox of sex and recombination. Nat. Rev. Genet. 3: 252–261.
Payseur B.A. and Nachman M.W. 2000. Microsatellite variation and recombination rate in the human genome. Genetics 156: 1285–1298.
Peck J.R. 1994. A ruby in the rubbish: Beneficial mutations and the evolution of sex. Genetics 137: 597–606.
Penn D.J. and Potts W.K. 1999. The evolution of mating preferences and major histocompatibility complex genes. Am. Nat. 153: 145–164.
Perez-Tome J.M. and Toro M. 1982. Competition of similar and non-similar genotypes. Nature 299: 153–154.
Perrot V., Richerd S., and Valero M. 1991. Transition from haploidy to diploidy. Nature 351: 315–317.
Poon A. and Chao L. 2003. Drift increases the advantage of sex in RNA bacteriophage phi6. Genetics 166: 19–24.
Provine W. 1986. Sewall Wright and evolutionary biology. University of Chicago Press, Chicago.
Queitsch C., Sangster T.A., and Lindquist S. 2002. Hsp90 as a capacitor of phenotypic variation. Nature 417: 618–624.
Rice W.R. 1996. Evolution of the Y sex chromosome in animals. Bioscience 46: 331–343.
Robertson A. 1966. A mathematical model of the culling process in dairy cattle. Anim. Prod. 8: 95–108.
Rutherford S.L. 2000. From genotype to phenotype: Buffering mechanisms and the storage of genetic information. BioEssays 22: 1095–1105.
Rutherford S.L. and Lindquist S. 1998. Hsp90 as a capacitor for morphological evolution. Nature 396: 336–342.
Schaaper R.M. 1998. Anitmutator mutants in bacteriophage T4 and Escherichia coli. Genetics 148: 1579–1585.
Skaletsky H., Kuroda-Kawaguchi T., Minx P.J., et al. 2003. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 423: 825–837.
Smith R.J., Kamiya T., and Horne D.J. 2006. Living males of the ‘ancient asexual’ Darwinulidae (Ostracoda: Crustacea). Proc. R. Soc. Lond. B 273: 1569–1578.
Sniegowski P.D., Gerrish P.J., and Lenski R.E. 1997. Evolution of high mutation rates in experimental populations of Escherichia coli. Nature 387: 703–705.
Sniegowski P., Gerrish P.J., and Johnson T. 2000. Evolution of mutation rates: Separating causes from consequences. BioEssays 22: 1057–1066.
Spencer C.C.A., Deloukas P., Hunt S., Mullikin J.C., Myers S., Silverman B., Donnelly P., Bentley D., and McVean G. 2006. The influence of recombination on human genetic diversity. PLoS Genet. 2: e148.
Stemmer W.P.C. 1994. Rapid evolution of a protein in vitro by DNA shuffling. Nature 370: 389–391.
Stephenson A.G. and Winsor J.A. 1986. Lotus corniculatus regulates offspring quality through selective fruit abortion. Evolution 40: 453–458.
Stern A., Brown M., Nickel P., and Meyer T.F. 1986. Opacity genes in Neisseria gonorrhoeae: Control of phase and antigenic variation. Cell 47: 61–71.
Tomlinson I.P.M., Novella I.S., and Bodmer W.F. 1996. The mutation rate and cancer. Proc. Natl. Acad. Sci. 93: 14800–14803.
van Ham S.M., van Alphen L., Mooi F.R., and van Putten J.P. 1993. Phase variation of H. influenzae fimbriae: Transcriptional control of two divergent genes through a variable combined promoter region. Cell 73: 1187–1196.
Vrijenhoek R.C. 1994. Unisexual fish: Model systems for studying ecology and evolution. Annu. Rev. Ecol. Syst. 25: 71–96.
Weiser J.N., Love J.M., and Moxon E.R. 1989. The molecular mechanism of phase variation of H. influenzae lipopolyaccharide. Cell 59: 657–665.
Weissman A. 1889. The significance of sexual reproduction in the theory of natural selection. In Essays upon heredity and kindred biological problems (ed. E.B. Poulton et al.), pp. 252–297. Clarendon Press, Oxford.
Welch D.B.M., Welch J.L.M., and Meselson M.S. 2008. Evidence for degenerate tetraploidy in bdelloid rotifers. Proc. Natl. Acad. Sci. 105: 5145–5149.
Welch D.M. and Meselson M. 2000. Evidence for the evolution of bdelloid rotifers without sexual reproduction or genetic exchange. Science 288: 1211–1215.
Welch J.L.M., Welch D.B.M., and Meselson M.S. 2004a. Cytogenetic evidence for asexual evolution of bdelloid rotifers. Proc. Natl. Acad. Sci. 101: 1618–1621.
Welch D.B.M., Cummings M.P., Hillis D.M., and Meselson M.S. 2004b. Divergent gene copies in the asexual class Bdelloidea (Rotifera) separated before the bdelloid radiation or within bdelloid families. Proc. Natl. Acad. Sci. 101: 1622–1625.
|



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}