|
Problem 23.1
An asexually reproducing organism can reduce its total rate of deleterious mutation, U, by devoting more resources to repairing damage to its DNA. However, this diversion of resources reduces its fitness by a factor (1 – exp(–2U)). (See Fig. P23.1.) (That is, if U were very large, fitness would not be reduced, but if U = 1 [say], it would be reduced by a factor 1 – exp(–1) = 0.632.)
|
| *i) |
What is the optimal mutation rate? What is the fitness at this optimum? HINT 23A
|
| **ii) |
Now consider a sexually reproducing organism. For simplicity, assume that it has ten pairs of chromosomes of equal size and that there is negligible recombination within each chromosome pair. What mutation rate will evolve? HINT 23B
|
|
Problem 23.2
A population of asexually reproducing bacteria has a genomic rate of mutation to deleterious alleles of U = 0.002 per generation; these alleles reduce fitness by s = 2%. Because the population must in the long run trace its ancestry back to the fittest class of mutation-free individuals (Fig. 19.19), we focus on the numbers of this class. NOTE 23A
|
| *i) |
What is the expected number of deleterious alleles carried by each individual? What fraction of the population is free of deleterious alleles? What is the average fitness, relative to this fittest genotype? HINT 23C
|
| *ii) |
If the population size is steady, the absolute mean fitness must be 1. What is the expected absolute fitness of a mutation-free individual?
|
| **iii) |
A mutator allele arises, which increases mutation rate by a factor 100 to an average of U* = 0.2 mutations per generation. Assume that this mutator allele arises in the fittest genotype and so initially is not associated with any deleterious mutations. On average, what fraction of its offspring will themselves carry no deleterious mutations?
|
| **iv) |
What is the expected number of copies of the mutator allele that are still free of any deleterious mutations after t generations? HINT 23D NOTE 23B
|
| **v) |
Summing over all future generations, how many copies of the mutator allele, in the fittest background, are expected to exist?
|
| **vi) |
Now suppose that favorable mutations arise at a very low rate μ = 10–10 per genome per generation and give a selective advantage s* = 1%. Such mutations can only fix if they arise on the fittest class, unencumbered by any deleterious alleles. What is the rate at which such favorable mutations become established in a population of 107 individuals? HINT 23E
|
| **vii) |
Assume that the mutator allele increases the mutation rate to favorable alleles by the same factor of U*/U = 100 as for deleterious alleles. Think of a single individual that carries the mutator allele. What is the chance that it, or its offspring, will acquire the favorable mutation, so that the mutator allele is swept up to high frequency? What is the chance that a mutator allele will be the lucky one that is swept to fixation, compared to the nonmutator alleles?
|
| ***viii) |
If it is produced in association with the mutator allele, what will happen to the new favorable allele? HINT 23F
|
|
Problem 23.3
A sexually reproducing population maintains a constant size, with equal proportions of males and females.
|
| *i) |
A parthenogenetic strain arises, with females who produce only daughters that are genetically identical to themselves. How rapidly will this strain increase?
|
| **ii) |
Suppose that males help rear the offspring. A female with no help from a male rears only one-fourth as many offspring. Initially, males mate randomly with both sexual and asexual females. What will happen? HINT 23G
|
|
Problem 23.4
A dominant allele arises in a hermaphroditic plant that causes it to self-fertilize.
|
| **i) |
What is the expected rate of increase from low frequency if selfing plants do not export pollen? HINT 23H
|
| **ii) |
What is the expected rate of increase from low frequency if selfing plants export pollen at the same rate as the wild type? ? HINT 23I
|
|
Problem 23.5
Beneficial alleles, which increase fitness by s = 5%, arise at a rate of 10–9 per genome per generation in a population of 109 asexual haploid microbes.
|
| *i) |
How long before one of these alleles becomes established and starts to increase?
|
| *ii) |
How long will it take before the allele reaches high frequency (say, 50%) in the population? HINT 23J
|
| *iii) |
During the time the first allele takes to fix, how many other alleles do we expect to increase?
|
| **iv) |
What is the rate of substitution of beneficial alleles in this asexual population? HINT 23K
|
| *v) |
What would be the rate with sexual reproduction?
|
| **vi) |
Would the relative advantage of sex be greater or less in a smaller population? HINT 23L
|
|
Problem 23.6
Diploid females produce a large number of offspring, which grow up together. Competition is intense, and so only a small fraction of these survive. An allele P increases competitive ability and is very rare in the population (p << 1).
|
| **i) |
What is its frequency among emerging individuals if the mother mates with one male? How fast will the allele increase? HINT 23M
|
| **ii) |
What is its frequency if females mated with two males? How fast will the allele increase?
|
|
Problem 23.7
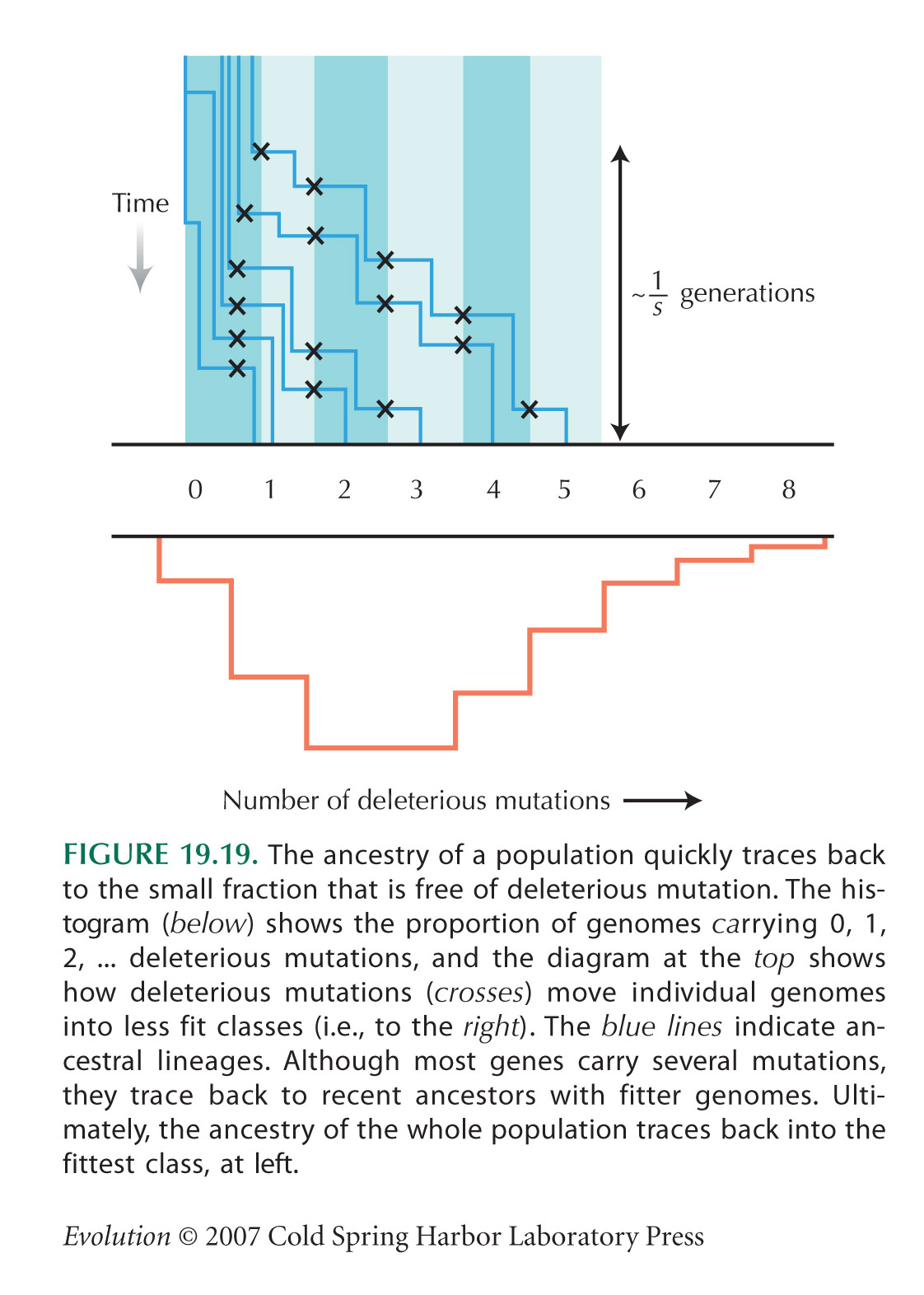
This problem explores the consequences of fluctuating environments, as illustrated in Figure 23.13. A haploid organism may grow up in small patches, each contributing a fixed number of individuals to a single breeding pool. NOTE 23C
Patches may be hot or cold, and they may be wet or dry; the relative proportions of the four possible environments (i.e., {cold, wet}, {hot, wet}, {cold, dry}, {cold, wet}) fluctuate from generation to generation. There are two unlinked genes: T, which influences adaptation to temperature, and M, which influences adaptation to moistness. In hot patches, the TH allele doubles survival relative to the alternative allele TC, but in cold patches, the TC allele has twice the survival of TH. Similarly, MW doubles survival relative to MD in wet patches but halves it in dry patches. The effects of the two genes multiply together, so that there is no epistasis within patches.
|
| **i) |
First, suppose that the environments are uncorrelated, so that there is no epistasis overall: Each of the four environments contributes 1/4 of the breeding population. What will happen if the TH allele is introduced into a population that is initially entirely TCMW? What will happen if the MD allele is then introduced? What is the final equilibrium? HINT 23N
|
| *ii) |
Will there be any selection on recombination at this equilibrium?
|
| ***iii) |
Now, suppose that wet patches are always cold, and dry patches are always hot. What will the equilibrium now be? HINT 23O
|
| **iv) |
What is the average fitness of the population? What does this tell you about whether selection favors increased or decreased recombination? HINT 23P
|
| ***v) |
Finally, suppose that the correlation between environments alternates from one generation to the next. There are always two kinds of patches, but in one generation, hot is always associated with wet and cold with dry, and in the next generation, this association reverses. Initially, the low recombination type is fixed, and all four genotypes are present. How does the average fitness of the population depend on the strength of linkage disequilibrium? HINT 23Q
|
|
Problem 23.8
This problem uses the infinitesimal model (p. 483) to explore the effect of stabilizing selection and recombination on quantitative variation. When a very large number of unlinked and additive alleles influence a trait, we can use a very simple approximation: When two individuals mate, their offspring have breeding values that follow a normal distribution, with the mean at the average of the parents, and a fixed within-family variance Vf, which is released by recombination. We ignore environmental variation throughout and just follow the breeding values directly. Measure variances relative to Vf throughout.
|
| *i) |
If the initial population has genetic variance Vg and individuals mate at random, what is the variance of the offspring population? HINT 23R
|
| *ii) |
What is the genetic variance at equilibrium?
|
| **iii) |
*C28* Stabilizing selection favors an optimum trait value z = 0, so that individual fitness is exp(–z2/2Vs). Here, Vs is a measure of the strength of stabilizing selection: Small Vs implies that a narrow range of trait values gives high fitness (p. 513). What is the mean fitness of a population with mean at the optimum and with genetic variance Vg? Assume that the population follows a normal distribution (see Chapter 28). HINT 23S
|
| **iv) |
After selection, what is the variance of the population? HINT 23T
|
| **v) |
What is the variance at equilibrium? HINT 23U
|
| *vi) |
Now, directional selection acts, with selection gradient β (p. 478). Thus, fitness is now given by exp(+βz – z2/2Vs), when the population mean is at z = 0. What is the change in the mean over the next generation?
|
| ***vii) |
What would happen if reproduction were asexual? HINT 23V NOTE 23D
|
|
Problem 23.9
This problem shows how inherited variation of fitness inflates the rate of random drift, which in turn interferes with selection (p. 677). If the variance of the number of offspring of a gene is v, then the variance of allele frequency increases at a rate (v/2N)pq, where there are 2N genes in the population (p. 419). NOTE 23E However, this assumes that fitnesses in successive generations are independent. If fitness is inherited, then random fluctuations in fitness will persist and have a larger effect. This point, first made by Robertson (1966), can be illustrated quite simply.
Suppose that there is a variance in fitness v caused by variation at unlinked genes. In a stable population, the average fitness is 1, but individual genes experience a fluctuation in fitness 1 + δ; the mean of δ is zero and its variance is v.
|
| **i) |
If a gene has, by chance, an expected fitness 1 + δ now, what will its expected fitness be in the next generation? What will it be in successive generations? HINT 23W
|
| **ii) |
What is the total expected number of copies left by a gene with fitness 1 + δ now? HINT 23X
|
| **iii) |
What is the variance in the total number of copies left, summed over future generations, in terms of the variance in immediate fitness, v? What is the rate of drift due to this inherited variation in fitness?
|
| **iv) |
Individual genes have a total variance in fitness of 1.25; the heritability of fitness is 20%, and the inherited component of fitness variation is due to unlinked genes. What is the effective population size, Ne, relative to actual numbers, N?
|
| *v) |
The probability of fixation of an allele with selective advantage s is approximately 2s(Ne/N). By how much does inherited variance in fitness reduce the chances of fixation of such an allele?
|
|
Problem 23.10
This problem shows how sexual reproduction can greatly reduce the mutation load, provided genes interact in a certain way (p. 680).
|
| *i) |
What is the mean fitness of an asexual population, which has a total mutation rate of U = 2 per genome, per generation? HINT 23Y
|
| *ii) |
What is the mean fitness of a sexually reproducing population, with the same total mutation rate, when the effect of different mutations multiply together?
|
| **iii) |
Now, suppose that the effect of deleterious mutations increases as more and more of them accumulate: The fitness of an individual with n deleterious mutations is W = exp(–(1/2)(n/200)4). (See Figure P23.3.) If the average individual has 200 deleterious mutations, what is the decrease in relative fitness (i.e., δ(W0/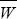 )) caused by one extra mutation? Given this selection coefficient and the total mutation rate U = 2, show that there are indeed 200 deleterious mutations on average. )) caused by one extra mutation? Given this selection coefficient and the total mutation rate U = 2, show that there are indeed 200 deleterious mutations on average.
|
| **iv) |
Sketch the distribution of the number of deleterious mutations. HINT 23Z
|
| *v) |
What is the mean fitness of the population? How does it compare with the mean fitness of an asexual population?
|
|
Problem 23.11
Several groups of insects, including the Hymenoptera (ants, bees, and wasps), are haplodiploid, that is, fertilized eggs develop as females, and unfertilized eggs develop as males.
|
| **i) |
How quickly will a dominant allele, A, that causes haplodiploidy increase? Females heterozygous for this allele produce unfertilized eggs that develop as sons; all their fertilized eggs develop as females. Haploid males carrying the new allele mate with wild-type females to produce half sons and half daughters, in the usual way. HINT 23AA
|
| **ii) |
Recessive lethal alleles reduce the fitness of heterozygotes by s = 2% and are fatal when present in a single copy in a haploid male. If the mutation rate to such recessive lethals is U = 0.2 per diploid genome per generation, then what is the average fitness of haploid males, immediately after the origin of haplodiploidy? HINT 23BB
|
| ***iii) |
What would the mean fitness be once the haplodiploid population reached equilibrium? HINT 23CC HINT 23DD
|
|
Problem 23.12
Why are there two sexes? That is, why do most eukaryotes produce either small male gametes or large female gametes? Imagine that a large zygote gives a greater chance of survival but that an individual can produce more gametes if these are smaller. In this problem, we will see whether this leads to the evolution of two sizes of gametes.
Assume that, initially, gametes are of size m and that individuals can produce n = A/m of these. Zygotes are of size 2m and survive with a probability proportional to (2m)k. We assume that k > 1, so that there are increasing returns to larger zygote size. The maximum possible size is M.
|
| *i) |
When there is only a single type in the population, will fitness increase or decrease with gamete size?
|
| **ii) |
Will producers of very small gametes be able to invade a population that produces large gametes? HINT 23EE
|
| **iii) |
What is final equilibrium? HINT 23FF
|
|
Problem 23.13
This problem looks at the evolution of dominance (pp. 690–691). Fitness W is proportional to the flux through a metabolic pathway, which depends on the concentration of an enzyme E and of a substrate X: W = EX/(E + X). There is also a cost to producing the enzyme, which reduces fitness by a factor exp(–βE).
|
| **i) |
What is the optimal enzyme concentration for a low cost, β = 0.01/X? For a high cost, β = 0.5/X? Sketch a graph of the optimal concentration against β. HINT 23GG
|
| **ii) |
Suppose that deleterious mutations eliminate gene function, so that in a heterozygote, the enzyme concentration is halved; the cost of production, exp(–βE), remains the same, however. Assuming an optimal enzyme concentration, what is the fitness of the heterozygote, relative to the homozygote? Consider cases of low cost and high cost, as in i). HINT 23HH
|
Robertson A. 1966. A mathematical model of the culling process in dairy cattle. Anim. Prod. 8: 95–108.



{kind=link}
{kind=link}