NOTE 19B: These data are from Bumpus (1899). Several reanalyses have been made, including those by Johnston et al. (1972), O’Donald (1973), and Schluter (1988).
NOTE 19C: These data are from Mallet et al. (1990).
NOTE 19D: The sequences also differ by insertions and deletions (indels), which appear as gaps in the alignment of the two sequences and which slightly alter the total number of sites that are compared. Because it is not clear how long insertion and deletion mutations are, we ignore these gaps in this question.
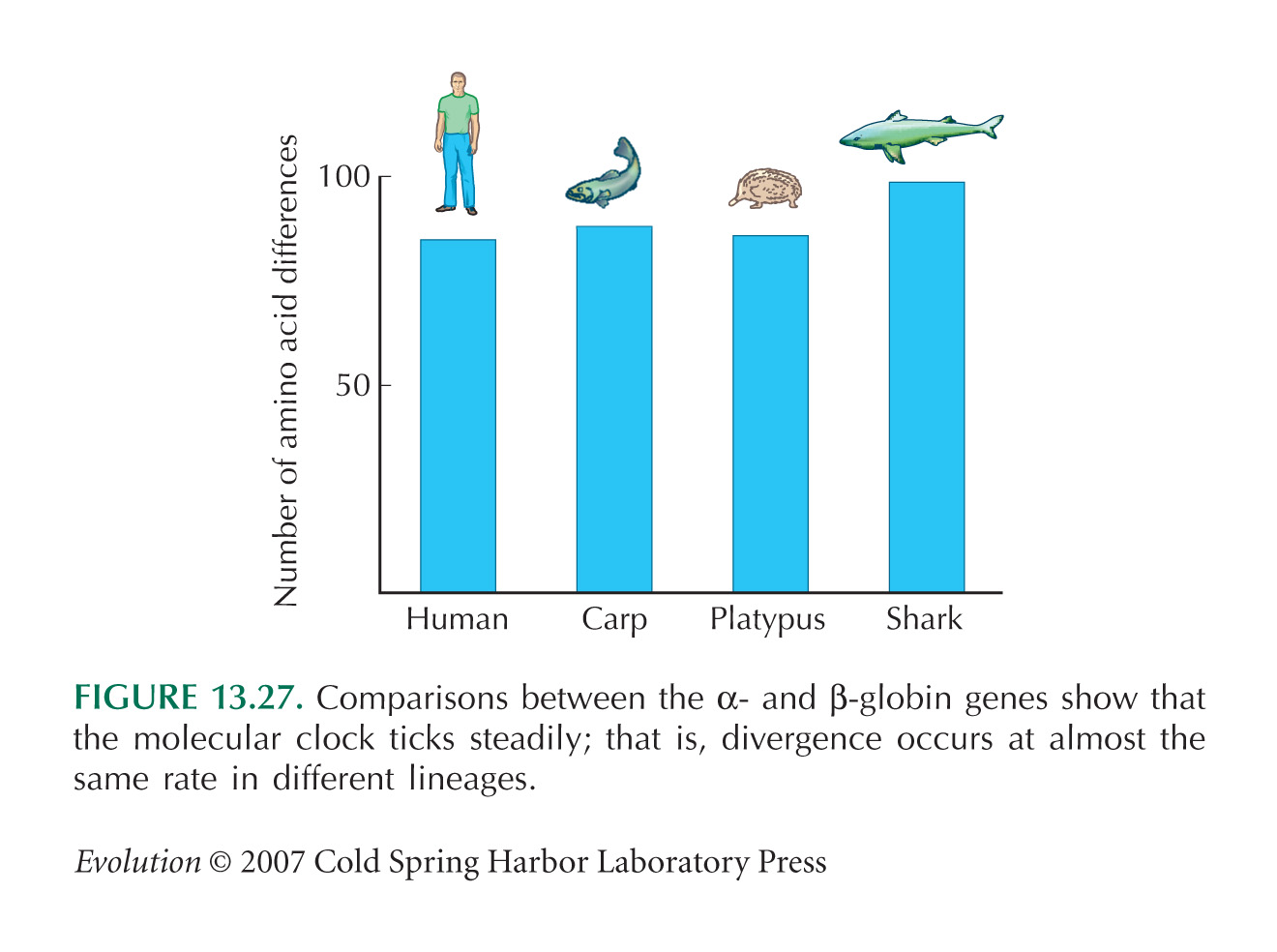
NOTE 19E: These data are from Table 4.5 of Kimura (1983). They are summarized in Figure 13.27.
NOTE 19F: This analysis is from Chapter 4 of Kimura (1983). Kimura uses a formula relating var(dij) to var(xi) in which n has been replaced by n + 1; it is unclear why.
NOTE 19G: The data are for 25 genes with Ps > 10, and excluding genes that were sequenced because they were thought to have been positively selected (line 3 of Table 1 of Smith and Eyre-Walker 2002). The estimates are slightly different because Smith and Eyre-Walker use a denominator of Ps + 1 to reduce bias; Box 19.1 gives the simpler formula with Ps in the denominator.
NOTE 19H: This example is courtesy of Darren Obbard, IEB, Edinburgh.
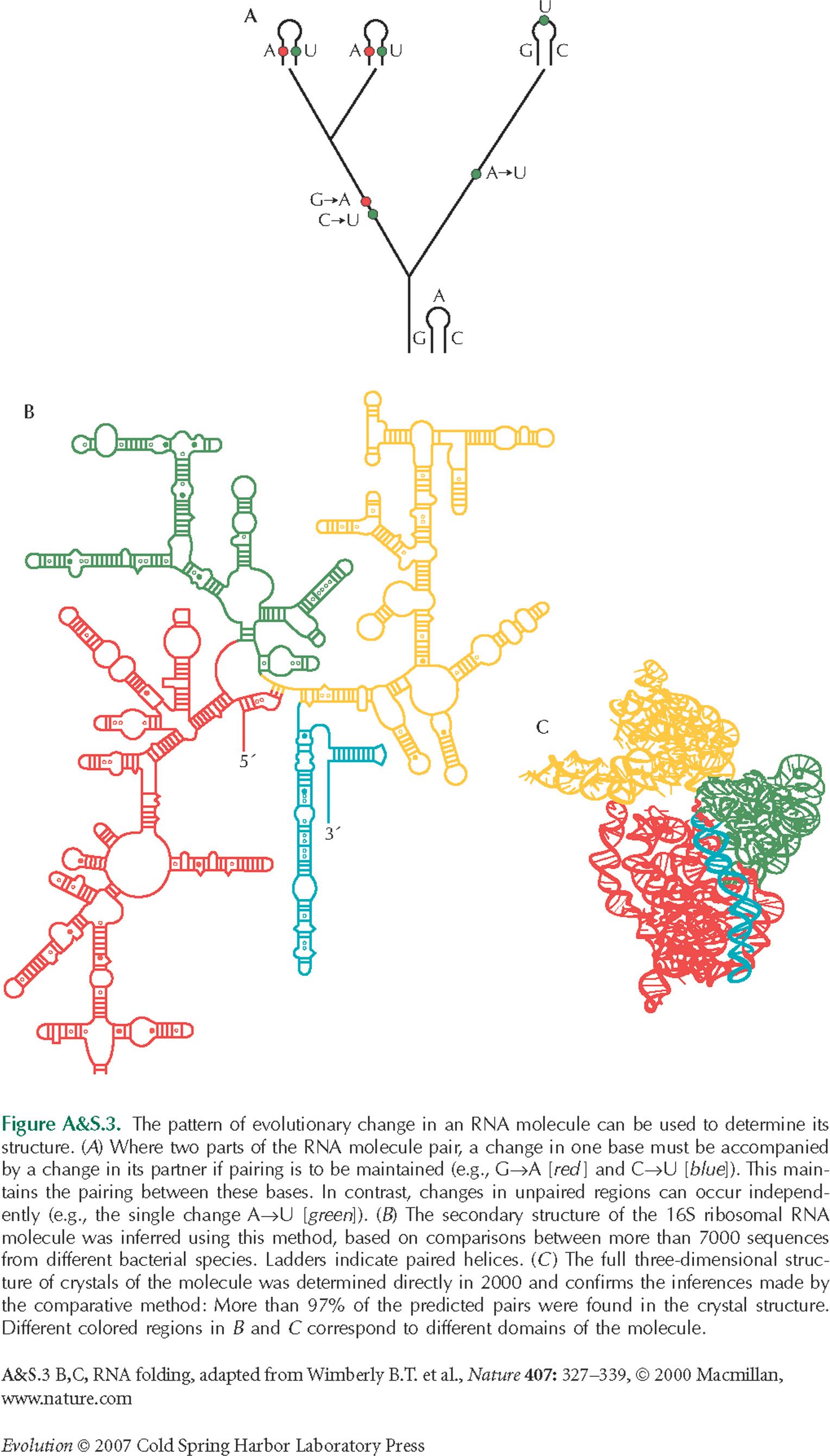
NOTE 19I: This pattern of correlated evolution can be used to infer the presence of pairing: Recall Figure A&S3.
NOTE 19J: This example is based on Innan and Stephan (2001).
NOTE 19K: The increase in a neutral allele, caused by its association with a selected allele, is know as hitchhiking (Maynard Smith and Haigh 1974).
NOTE 19L: This problem follows the derivation in Maynard Smith and Haigh (1974). It follows allele frequencies, whereas Problem 19.9 gets the same result by following coalescence and recombination of ancestral lineages.
{kind=link}
{kind=link}