Chapter 3 Notes
Evidence for Evolution
Introduction
The well-known statue shown in the opening figure was cast by Wolfgang Hugo Rheinhold in 1895. Vladimir Lenin kept a copy on his desk.
A translation of Pope John Paul II’s 1996 message to the Pontifical Academy of Sciences was printed in the Quarterly Review of Biology (72: 381–406). It includes a clear explanation of the nature of scientific theory and the reasons why science need not conflict with religious belief. The Quarterly Review of Biology also published four commentaries; Ruse’s article in particular gives some useful historical background.
Christian theologians reconcile religious beliefs with natural law in a variety of ways. The regularity of Newton’s laws stimulated the development of Deism in the 18th century. This holds that God created the world and the laws that govern it but then played no further part in its operation. (God is seen as the watchmaker and the Universe as the watch.) Theistic evolution (a view that can be traced back to Thomas Aquinas) sees God as the primary cause of all that happens, but acting through a chain of “secondary causes,” which include the laws of physics and of biological evolution. McGrath (1999, p. 102) gives an introduction to these arguments.
There is an extensive literature on how theology can be reconciled with evolution by natural selection. For a discussion from the point of view of a philosopher of biology, see Ruse (2001). McGrath (1999) gives a balanced overview from a theological point of view. Witham (2002) provides a readable account of the conflict between biologists and fundamentalist Christians in the United States; the science is described somewhat inaccurately, but this is an excellent overview of the various religious and political views.
Peters and Bennett (2003) include several articles on the response of non-Christian religions to evolutionary ideas. Sakura (1998) discusses the reception of Darwinism in Japan.
Evidence for Evolution
Patterns of Relationship Provide the Most Powerful Evidence for Evolution
Universally Shared Features
Mutations to stop codons, which cause premature termination of translation, are known as nonsense or amber mutations. It is remarkable that tRNA mutations can suppress nonsense mutations, because this causes many other proteins to be extended. There are several solutions to this puzzle.
-
Short extensions to proteins somewhat reduce growth, but premature termination of an essential protein is lethal.
-
There are three stop codons; if the nonsense mutation is to a stop codon that is rarely used, then suppressing recognition of that codon may cause little harm.
-
The tRNAs coded by suppressor mutations only sometimes recognize stop codons. Thus, most proteins terminate correctly. For more details, see Watson et al. (2004, pp. 471–475).
Hierarchical Classification
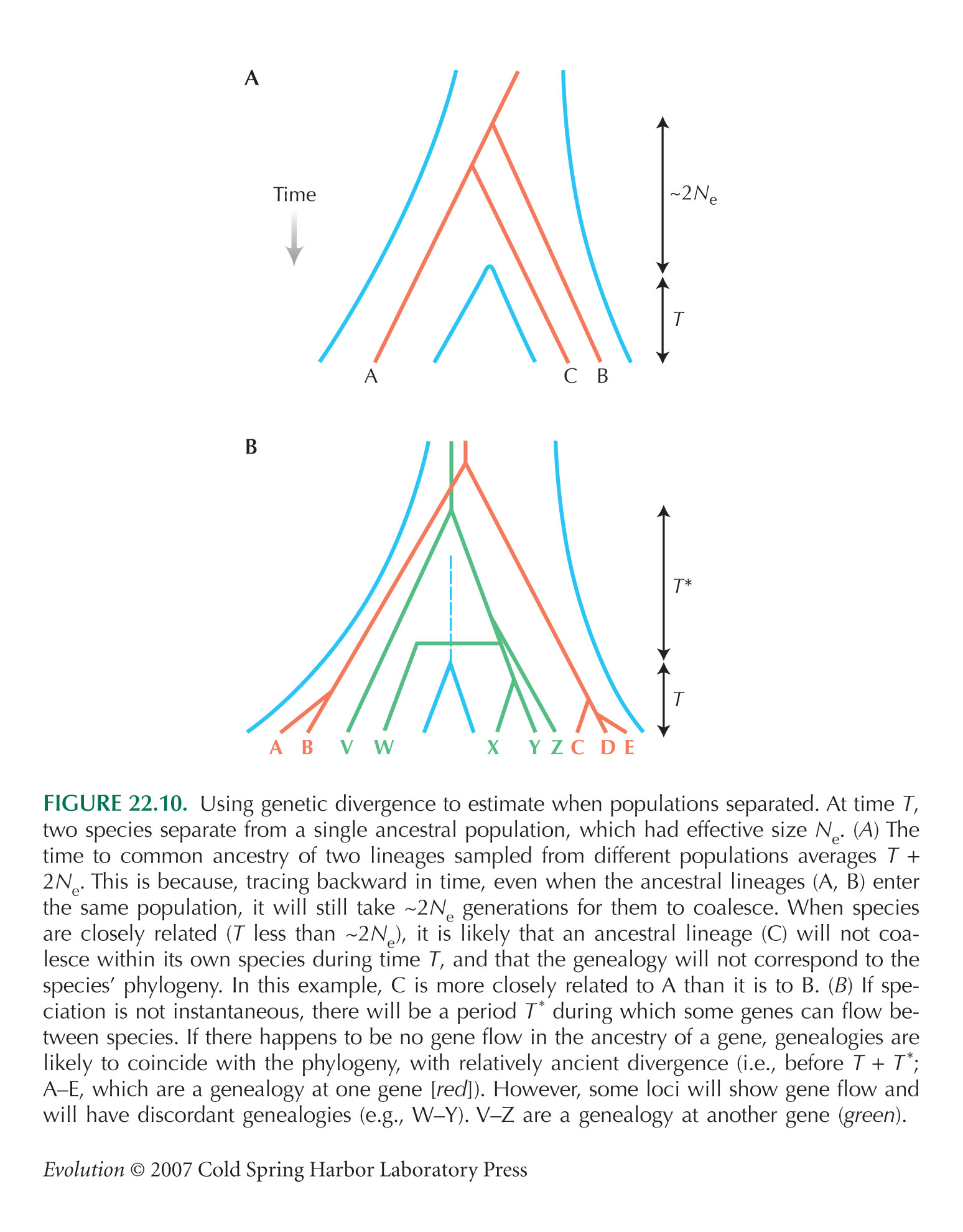
Some apparent exceptions provide further support for evolution. Closely related species may share polymorphisms that are inherited from their ancestral population. In such cases, genes from different species can be more similar to each other than genes from within the same species (see Fig. 22.10). For example, humans share several polymorphic types of gene at the major histocompatibility complex, implying that these types were all present in our common ancestor with chimpanzees (see pp. 541 and 628). This discordance between genealogy and phylogeny is known as lineage sorting.
Eldredge (2005, p. 236) shows how the classification of musical instruments differs from a biological phylogeny, with new features incorporated into otherwise unrelated groups. Such lateral transfers are also seen in the classification of languages (pp. 752–754). In bacteria and archaea, very rare transfers of genes between divergent lineages also make it impossible to classify them into a strictly branching phylogeny (pp. 182–191). In all of these examples, “descent with modification” is combined with processes of lateral transfer in a well-understood way, making classification more difficult than for the groups studied by Linnaeus, Darwin, and most biologists since. We discuss these issues in more detail in Chapters 15, 22, and 27.
Geographic Distribution
Eldredge (2005, p. 64) describes how Darwin’s observations of mockingbirds, tortoises, and finches on the Galápagos led him to the theory of evolution. See also On the Origin of Species, Chapters 11 and 12.
Evolutionary Processes Can Be Observed Directly
Figure 3.9 is from Molla et al. (1996); see also Condra et al. (1996).
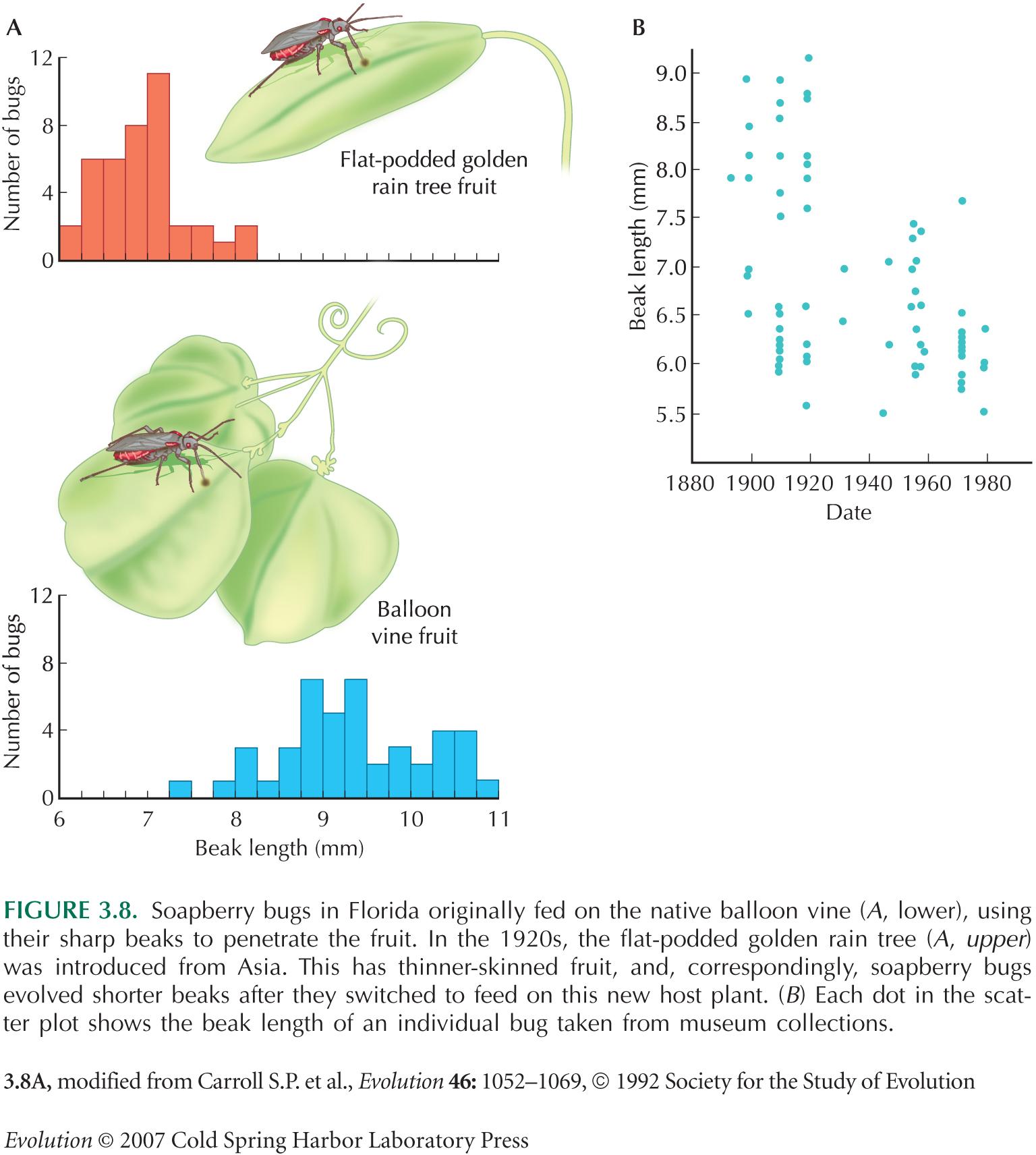
The rapid evolution of soapberry bugs in Florida (Fig. 3.8) is described by Carroll and Boyd (1992).
The Fossil Record Provides Several Lines of Evidence for Evolution
Darwin vividly describes his fossil finds in South America in his account of the voyage of the Beagle (1839).
Natural Selection Causes The Appearance of Design
The quote on page 75 is from Chapter 13 of On the Origin of Species.
Jacques Monod explored the idea of “evolution by tinkering” in his book, Chance and Necessity (1971).
Objections to Evolution
Objections to the Fact of Evolution
Continental drift can now be observed directly, by accurate laser interferometry. However, it was established first by indirect arguments.
The evolution of whales from land mammals is discussed by Thewissen et al. (2001) and by Gatesy and O’Leary (2001).
Objections to Natural Selection as the Cause of Adaptation
In a letter to Charles Lyell (10th December, 1859), Darwin wrote:
I have heard by round about channel that Herschel says my Book “is the law of higgledy-pigglety”. What this exactly means I do not know, but it is evidently very contemptuous.
The “argument from design” is discussed in detail, from an evolutionary perspective, by Ruse (2003). For a philosopher’s view, see Sober (2005).
Behe (1996) introduced the term “irreducible complexity” to recent discussions of “intelligent design.” However, the idea that complex adaptations consist of many interdependent parts was clearly stated by William Paley in 1802. The basic arguments showing how complex structures may evolve were given by Darwin in the On the Origin of Species. Ruse (2003, Chapter 15) refutes the “intelligent design” arguments, whereas Young and Edis (2004) deal with the scientific examples, such as the bacterial flagellum; see also Liu and Ochman (2007). Behe (2007) has revised his arguments, placing more emphasis on the improbability of multiple mutations. See Dawkins (2007) and Miller (2007). For more sources, see Chapter 3, Further Reading.
Pinker (1997, 2002) and Buller (2005) discuss the evolution of human nature from different points of view. These are just three examples from a burgeoning literature.
Eldredge and Gould (1972) first proposed the theory of punctuated equilibrium. Charlesworth et al. (1982) provide a critical review, emphasizing that the observed pattern of punctuated equilibrium is consistent with the Evolutionary Synthesis. See also Eldredge (2005).
Assume that the human coding sequence consists of about 30,000 genes, of average length 1340 bp (Lander et al. 2001) and that about the same amount of noncoding sequence is functionally constrained. Each base has 4 = 22 states and so codes for 2 bits of information. Hence, the human genome encodes approximately (2 × 2 × 1340 × 30,000 bits). One byte is defined as 8 bits, and so this is approximately 20 Mb. (We ignore the redundancy of the code here, and the number of genes in the human genome is now thought to be lower than 30,000. So, this is an overestimate. On the other hand, more noncoding DNA may be functional than we have assumed here.)
The figure of 40,000 amino acid differences between humans and chimpanzees is from Mikkelsen et al. (2005), made by direct comparison of genome sequences. The figure of 1/3 for the fraction selected is from Fay et al. (2001). It is lower than expected from the estimates given on p. 536, which were made by Fay et al. (2001), based on divergence along the human lineage since we separated from old-world monkeys, 30 Mya. They estimated 150,000 adaptive differences (~35% of the total) or 15,000 total per Myr. So, we would expect (2 × 6 × 15,000) = 180,000 differences between chimpanzees and humans, who diverged about 6 Mya. Fay et al. (2001) give a slightly higher estimate of the total coding genome (50 Mb), possibly because they use a higher estimate of gene number.
For a detailed account of the complexity of the human brain, see Koch (2007).
For more on the Fisher quotation in Figure 3.19, see Chapter 17 Web Notes.
Science and Society
The Fact of Evolution Is Explained by Evolutionary Theory
Futuyma (1995) gives a clear explanation of what is meant by a scientific theory. See also the 2005 judgment against the Dover Area School District and its Board of Directors.
Ruse (2001, pp. 91 and 122) discusses why a “God of the Gaps” is seen as unsatisfactory; see also the commentaries accompanying Pope John Paul II’s article (discussed above).
For a recent view of the teaching of evolution in the United States, see Scott and Matzke (2007).
Antonovics et al. (2007) document reluctance in the medical literature to describe antibiotic resistance as “evolving”; they also show that whether press reports use “evolving” is strongly correlated with usage in the initial paper. This suggests that this may make “evolution” less prominent in general than it should be.
Understanding Nature and Humanity
The quote from the Louisiana Senate hearings is from Witham (2002, Chapter 14).
Gould (2001) sets out the case that science and religion have separate domains of authority.
Darwin used almost the same words to close On the Origin of Species, published 17 years later.
The phrase “nature red in tooth and claw” is from Tennyson’s In Memoriam (1850).
Further Reading
The National Academy of Sciences (U.S.A.) has recently updated its booklet “Science, Evolution and Creationism”; its website gives this and other resources for education about evolution. For further links, see Web Resources.
References
Antonovics J., Abbate J.L., Baker C.H., Faley D., Hood M.E., Jenkins C.E., Johnson L.J., Murray J.J., Panjeti V., Rudolf V.H.W., Sloan D., and Vondrasek J. 2007. Evolution by any other name: Antibiotic resistance and avoidance of the E-word. PLoS Biol. 5: 137–140.
Behe M. 1996. Darwin’s black box: The biochemical challenge to Darwinism. Free Press, New York.
Behe M.J. 2007. The edge of evolution: The search for the limits of Darwinism. Free Press, New York.
Buller D.J. 2005. Adapting minds. MIT Press, Cambridge, Massachusetts.
Carroll S.P. and Boyd C. 1992. Host race radiation in the soapberry bug: Natural history with history. Evolution 46: 1052–1069.
Charlesworth B., Lande R., and Slatkin M. 1982. A neo-Darwinian commentary on macroevolution. Evolution 36: 474–498.
Condra J.H., Holder D.J., Schleif W.A., Blahy O.M., Danovich R.M., Gabryelski L.J., Graham D.J., Laird D., Quintero J.C., Rhodes A., Robbins H.L., Roth E., Shivaprakash M., Yang T., Chodakewitz J.A., Deutsch P.J., Leavitt R.Y., Massari F.E., Mellors J.W., Squires K.E., Steigbigel R.T., Teppler H., and Emini E.A. 1996. Genetic correlates of in vivo viral resistance to indinavir, a human immunodeficiency virus type 1 protease inhibitor. J. Virol. 70: 8270–8276.
Darwin C. 1839. Journal of researches into the geology and natural history of the various countries visited by H.M.S. Beagle, under the command of Captain FitzRoy, R.N., from 1832 to 1836. Henry Colburn, London.
Dawkins R. 2007. “The Edge of Evolution” by Michael J. Behe: Inferior design. New York Times Sunday Book Review, July 1, 2007.
de Muizon C. 2001. Walking with whales. Nature 413: 259–260.
Eldredge N. 2005. Darwin: Discovering the tree of life. W.W. Norton, New York.
Eldredge N. and Gould S.J. 1972. Punctuated equilibria: An alternative to phyletic gradualism. In Models in paleobiology (ed. T.J.M. Schopf), pp. 82–115. Freeman, Cooper & Co., San Francisco, California. (Reprinted 1985 in Eldredge N. Time frames. Princeton University Press.)
Fay J., Wycoff G.J., and Wu C.I. 2001. Positive and negative selection on the human genome. Genetics 158: 1227–1234.
Futuyma D.J. 1995. Science on trial: The case for evolution. Sinauer Associates, Sunderland, Massachusetts.
Gatesy J. and O’Leary M.A. 2001. Deciphering whale origins with molecules and fossils. Trends Ecol. Evol. 16: 562–571.
Gould S.J. 2001. Rocks of ages. Jonathan Cape, London.
Koch C. 2007. The quest for consciousness. Rogers and Company, Englewood, California.
Lander E.S., Linton L.M., Birren B., Nusbaum C., Zody M.C., et al.; International Human Genome Sequencing Consortium. 2001. Initial sequencing and analysis of the human genome. Nature 409: 860–921.
Liu R.Y. and Ochman H. 2007. Stepwise formation of the bacterial flagellar system. Proc. Natl. Acad. Sci. 104: 7116–7121.
McGrath A.E. 1999. Science and religion: An introduction. Blackwell, Oxford.
Mikkelsen T.S., Hillier L.W., Eichler E.E., Zody M.C., Jaffe D.B., et al. The Chimpanzee Sequencing and Analysis Consortium. 2005. Initial sequence of the chimpanzee genome and comparison with the human genome. Nature 437: 69–87.
Miller K.R. 2007. Falling over the edge (review of The Edge of Evolution by Michael J. Behe). Nature 447: 1055–1056.
Molla A., Korneyeva M., Gao Q., Vasavanonda S., Schipper P.J., Mo H.M., Markowitz M., Chernyavskiy T., Niu P., Lyons N., Hsu A., Granneman G.R., Ho D.D., Boucher C.A., Leonard J.M., Norbeck D.W., and Kempf D.J. 1996. Ordered accumulation of mutations in HIV protease confers resistance to ritonavir. Nat. Med. 2: 760–766.
Monod J. 1971. Chance and necessity: An essay on the natural philosophy of modern biology. Alfred A. Knopf, New York.
Paley W. 1802. Natural theology. Reprinted 2006 (with an introduction and notes by M.D. Eddy and D.M. Knight). Oxford University Press, Oxford.
Penny D., Foulds L.R., and Hendy M.D. 1982. Testing the theory of evolution by comparing phylogenetic trees constructed from five different protein sequences. Nature 297: 197–200.
Peters T. and Bennett G. 2003. Bridging science and religion. Fortress Press, Minneapolis, Minnesota.
Pinker S. 2002. The blank slate: The modern denial of human nature. Viking Penguin, New York.
Pinker S. 1997. How the mind works. W.W. Norton, New York.
Pope John Paul II. 1996. Message to the Pontifical Academy of Sciences. Reprinted in Q. Rev. Biol. 72: 381–406, together with four commentaries.
Ruse M. 2003. Darwin and design. Harvard University Press, Cambridge, Massachusetts.
Ruse M. 2001. Can a Darwinian be a Christian? The relationship between science and religion. Cambridge University Press, Cambridge.
Sakura O. 1998. Similarities and varieties: A brief sketch on the reception of Darwinism and sociobiology in Japan. Biol. Philos. 13: 341–357.
Scott E.C. and Matzke N.J. 2007. Biological design in science classrooms. Proc. Natl. Acad. Sci. 104: 8669–8676.
Sober E. 2005. The design argument. In The Blackwell guide to the philosophy of religion (ed. W.E. Mann), pp. 117–147. Blackwell, Oxford.
Thewissen J.G.M., Williams E.M., Roe L.J., and Hussain S.T. 2001. Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls. Nature 413: 277–281.
Watson J.D., Baker T.A., Bell S.P., Gann A., Levine M., and Losick R. 2004. Molecular biology of the gene, 5th ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Witham L.A. 2002. Where Darwin meets the bible. Oxford University Press, Oxford.
Young M. and Edis T., eds. 2004. Why intelligent design fails. Rutgers University Press, New Jersey.
|



{kind=link}
{kind=link}
{kind=link}
{kind=link}