Chapter 20 Notes
Phenotypic Evolution
Evolutionary Optimization
Much of Evolution Can Be Understood as Optimization
Williams (1992) provides an excellent commentary on Paley’s arguments, together with extracts from Paley’s (1802) Natural Theology.
Box 20.1 The Logic of Optimization Arguments
The dung fly example is from Parker and Simmons (1994).
The example of metabolic networks in Escherichia coli comes from Ibarra et al. (2002); Edwards and Palsson (2000) explain the metabolic model.
The Optimum Is Defined Relative to Constraints on What Is Possible
Gould and Lewontin (1979) gave an influential critique of the adaptationist program. Parker and Maynard Smith (1990) clarify how optimization arguments can be used properly.
is an example of spandrels, which are used in the adaptationist argument by Gould and Lewontin (1979).
We have varied expectations as to how finely adjusted different features of organisms will be; this issue is discussed .
There Are Trade-offs between Components of Fitness
The optimal size of the genetic alphabet is discussed by Szathmary (1991) and in a more sophisticated argument by Szathmary (1992). The set of 6 base pairs was from Piccirilli et al. (1990). Szathmary (2004) gives an up-to-date review.
The actual code is “almost” the most efficient, in that Szathmary found that only one alternative combination of bases was arguably more efficient.
Trade-offs can be investigated by observing genetic correlations or by artificial selection experiments. We discuss these methods .
There has been extensive work on the adaptation of E. coli to changes in temperature; surprising, this does not seem to involve strong trade-offs. See Bennett et al. (1992) and subsequent papers in that series.
Aging
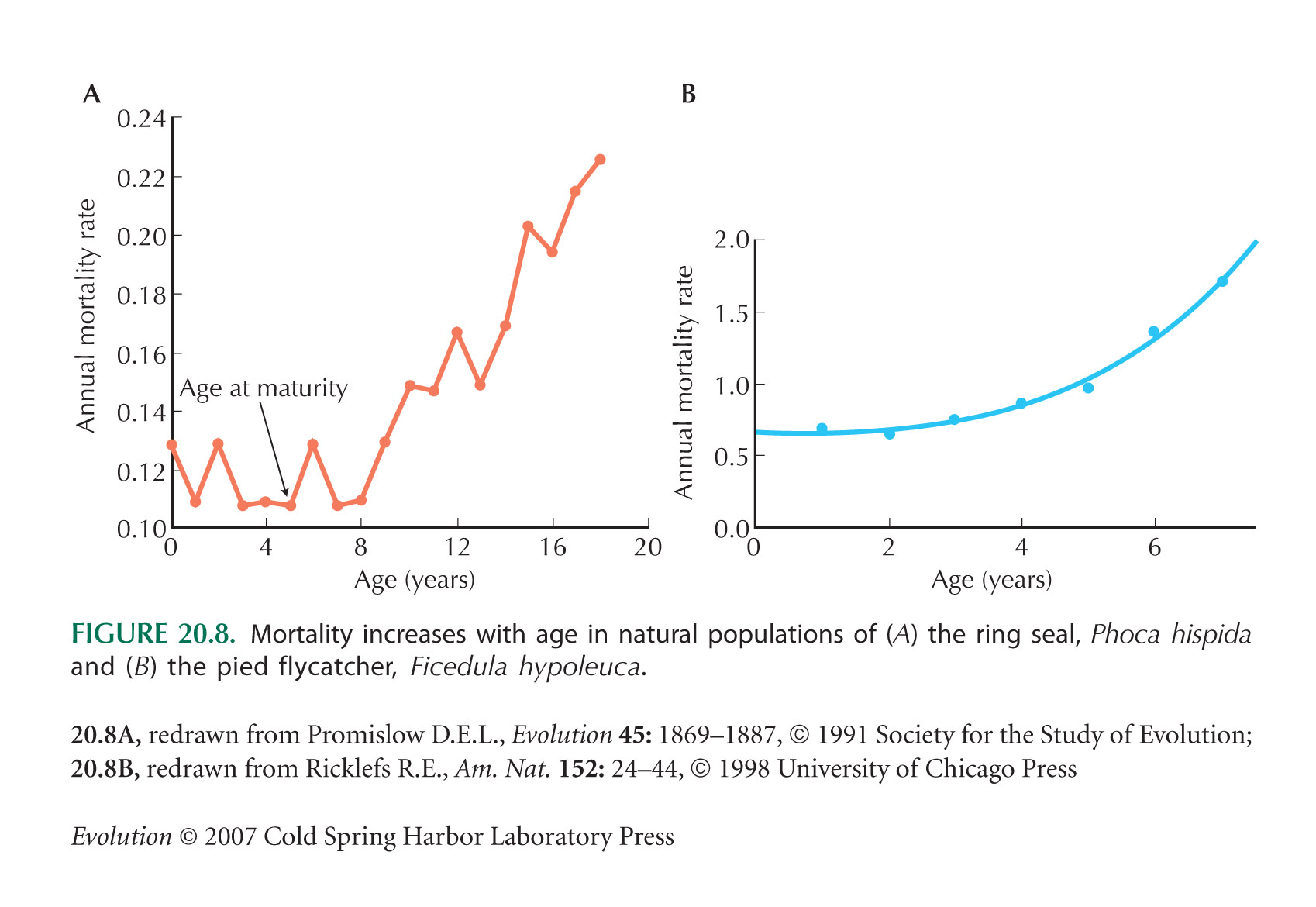
Aging Is Not Inevitable
The mortality data in Figure 20.8 are from Promislow (1991) and Ricklefs (1998).
Stewart et al. (2005) demonstrate senescence in the bacterium E. coli, which divides asymmetrically.
Aging Evolves Because the Old Make Little Contribution to Fitness
Fisher (1930) identified the weakened selection on later age, an argument from which Medawar (1946) identified the evolutionary explanation of aging. Hamilton (1966) gave the first mathematical analysis of how selection pressures change with age and set out the evolutionary explanation for senescence in detail. Charlesworth (2000) gives a history of these ideas.
Organisms that suffer less predation will also show less senescence, because survival and reproduction later in life will then be maintained by selection. One piece of evidence in favor of this evolutionary explanation is that birds show lower mortality than do mammals of similar size. Williams (1992) has another explanation in his book (p. 151): Birds typically become sexually mature long after they stop growing, whereas mammals start reproduction just before becoming fully grown. So, birds have to maintain their bodies for a long period before reproduction, and they just carry on working after that. These explanations are not incompatible. The first theory describes the selection regime that has been altered as a result of lower predation, and the other theory describes the physiological basis of the trade-off that has been reset in response (L. Partridge, pers. comm.). (A further complication in this example is that birds have been selected for small size, to enable flight, and so may tend to have a life history typical of mammals of larger size, including a longer life span.)
The example from social insects is from Keller and Genoud (1997).
Other examples of the loss of unselected functions include the evolution of host preferences in checkerspot butterflies (Euphydras editha) in response to loss of their usual food plant. Singer et al. (1993) show that this has led to an increase in preference for the new plant and a corresponding loss of preference for the original host. We see how changes in host preference can lead to speciation on pages 653–654.
Box 20.2 Selection Acts More Weakly on Later Life
This example comes from Roper et al. (1993); it is an extension of ideas presented in Chapter 17 (see ).
Aging May Evolve as Part of an Optimal Life History
The effects of genetic or experimental sterilization of Drosophila are reviewed by Partridge and Barton (1993).
The effects of manipulating egg number are reviewed by Godfray et al. (1991).
The effects of castration on wild Soay sheep is described by Jewell (1997). For estimates of survival rate, see Catchpole et al. (2000).
The Mutation Load May Be Concentrated on the Old
Haldane (1941) suggested that modifiers that postpone the age of onset of deleterious mutations might accumulate, leading to senescence.
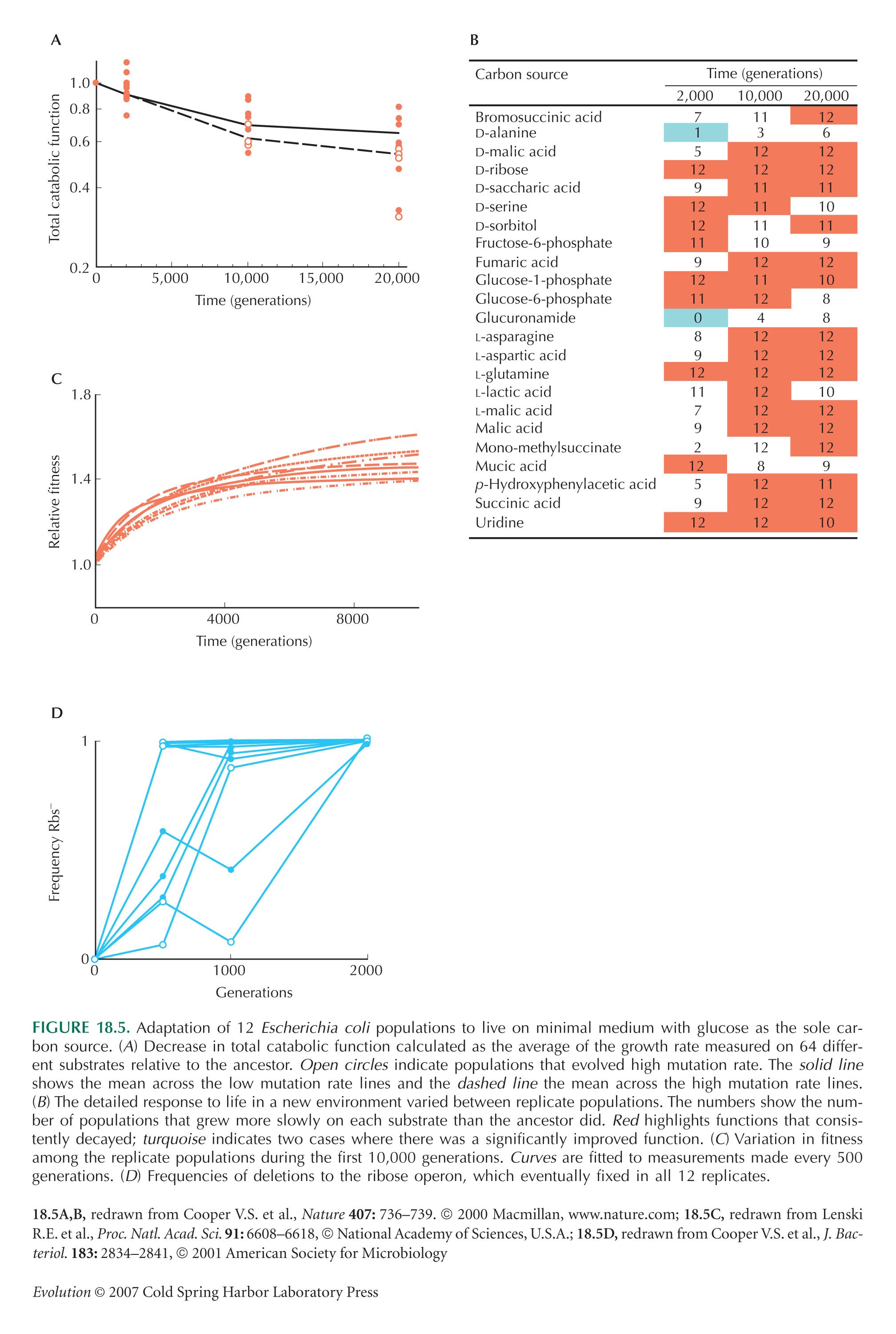
Cooper and Lenski (2000) demonstrate the loss of catabolic functions in replicate E. coli lines grown on minimal medium (see Fig. 18.5). This is an example where trade-offs can be distinguished from mutation accumulation. Selection positively favored deletions that caused loss of function, indicating that a new optimal life history is being approached through the trade-off between ability to metabolize different energy sources. This is not an instance of increased load from deleterious mutations, because the deletions are positively selected.
Hughes et al. (2002) observed an increase in additive variance in late life, which they took as evidence for mutation accumulation as a cause of senescence. However, other observations of how genetic variance and inbreeding depression change through time have given mixed results. See Snoke and Promislow (2003).
Aging Is Influenced by Conserved Mechanisms for Optimizing the Life History
Conserved mechanisms that regulate aging are explained . For a readable account of work on aging in yeast, see Guarente (2002). For recent reviews, see Guarente (2001) and Partridge and Gems (2002).
Evolutionary Games
An Evolutionarily Stable Strategy Cannot Be Displaced by Any Alternative
W.D. Hamilton (1966) introduced the idea of an evolutionarily stable strategy (ESS), but called it an “unbeatable strategy.”
Further details of the hawk–dove game are given .
Competition between Viruses Is an Evolutionary Game
The DIV (defective interfering virus) example of Figure 20.15 is from Nee and Maynard Smith (1990).
The Prisoner’s Dilemma game has been illustrated by an elegant with bacteriophage φ6.
The term “tragedy of the commons” is from Hardin (1968).
If There Is No ESS, Populations May Cycle: The Rock–Paper–Scissors Game
Strictly speaking, in the E. coli example, strategy S is an ESS, because C can only displace it above a critical threshold frequency, at which the poison becomes effective. However, this threshold is very low.
The different outcome in the E. coli experiment with an agar plate is due to the localization of interactions, rather than the growing conditions. If the transfer between agar plates is made by pressing the velvet pad several times in different orientations, the R strain fixes, just as in a stirred flask.
The E. coli bacteriocin example is from Kerr et al. (2002); see also Kirkup and Riley (2004).
Behavioral Polymorphism in Lizards Gives a Natural Example of the Rock–Paper–Scissors Game
In the Uta example, note that there are intermediate morphs that correspond to heterozygotes and that there is also a female polymorphism probably caused by the same locus. The cycling of males and females is coupled, and models suggest that the male cycles are speeded up by coupling with a female two-year cycle. See Sinervo (1996, 2001).
Sexual Selection
Sexual Selection Can Be Maladaptive
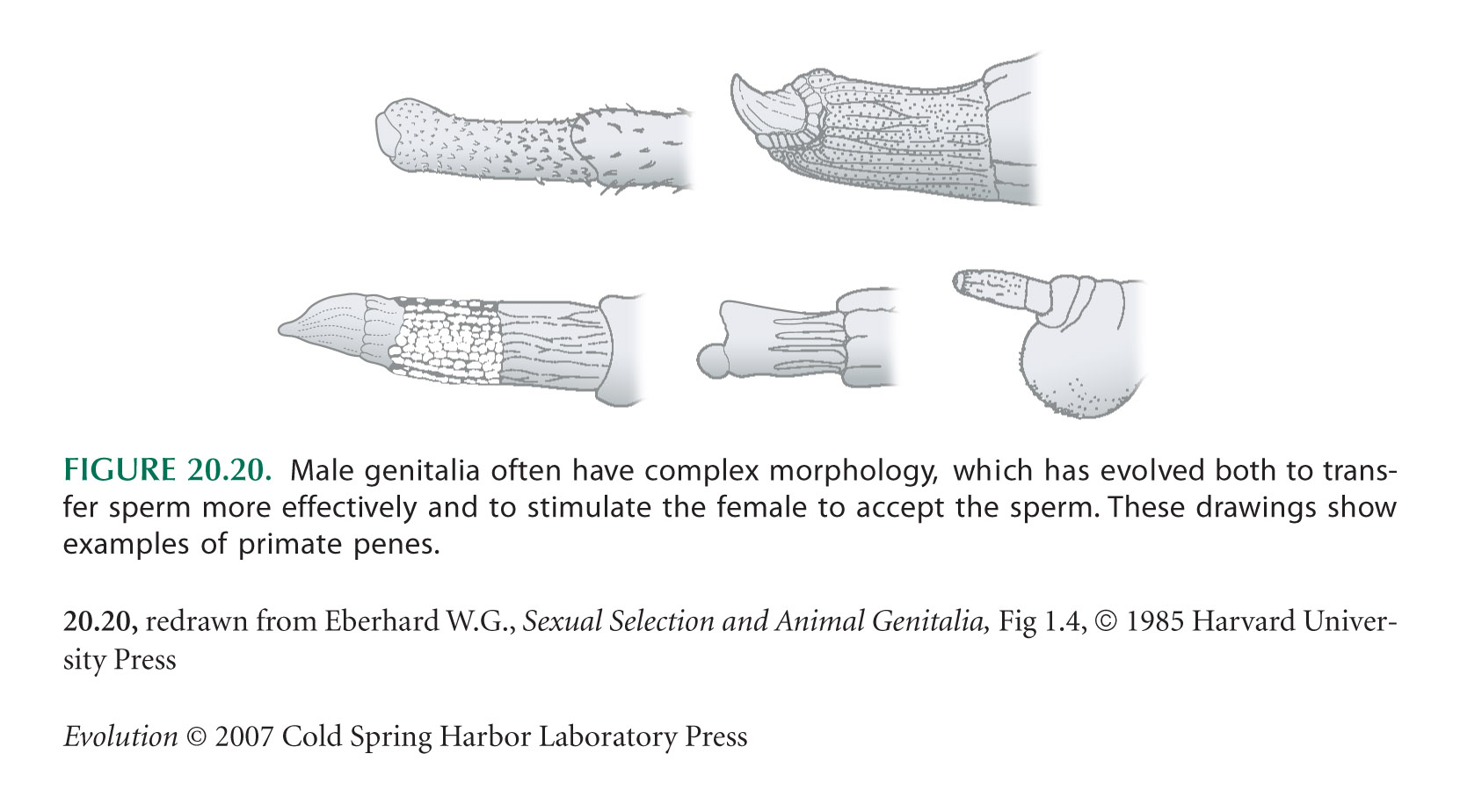
Eberhard (1986) gives a fascinating account of how sexual selection has led to extraordinary variation in animal genitalia (e.g., see Figs. 20.20 and 20.26).
The stickleback example of Figure 20.22 is from Hagen and Moodie (1979), and Hagen et al. (1980).
Tuttle and Ryan (1981) give a striking example of the costs of frog calls: Calling males are eaten by bats.
Sexual Selection Acts More Intensely among Males than among Females
Clutton-Brock (1988) reviews evidence on the variance in fitness in the two sexes; see also Merila and Sheldon (2000).
Clark (2002) reviews work on sperm competition in Drosophila and discusses how it may maintain variation.
Figures for fitness variance in red deer are from Kruuk et al. (2000).
Clark et al. (1995) show between genotypes of Drosophila melanogaster in sperm offense and defense. They also report an association between accessory protein alleles and sperm defense ability.
The quote is from Darwin (1882, p. 224). In this section, Darwin asks why the struggle is “between males for the possession of females” (to use his description of sexual selection in On the Origin), but does not clearly state the modern explanation.
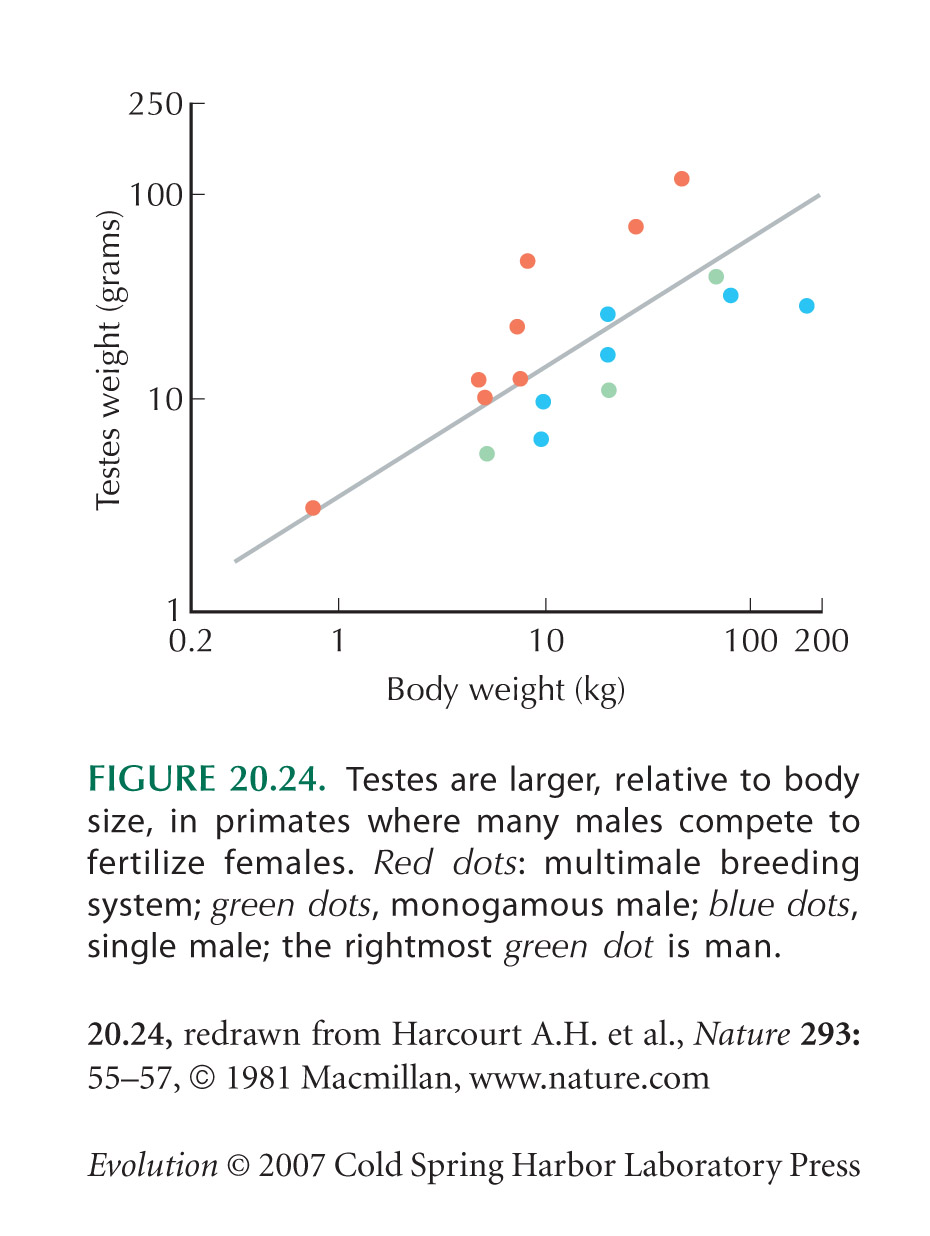
Figure 20.24 is based on Harcourt et al. (1981).
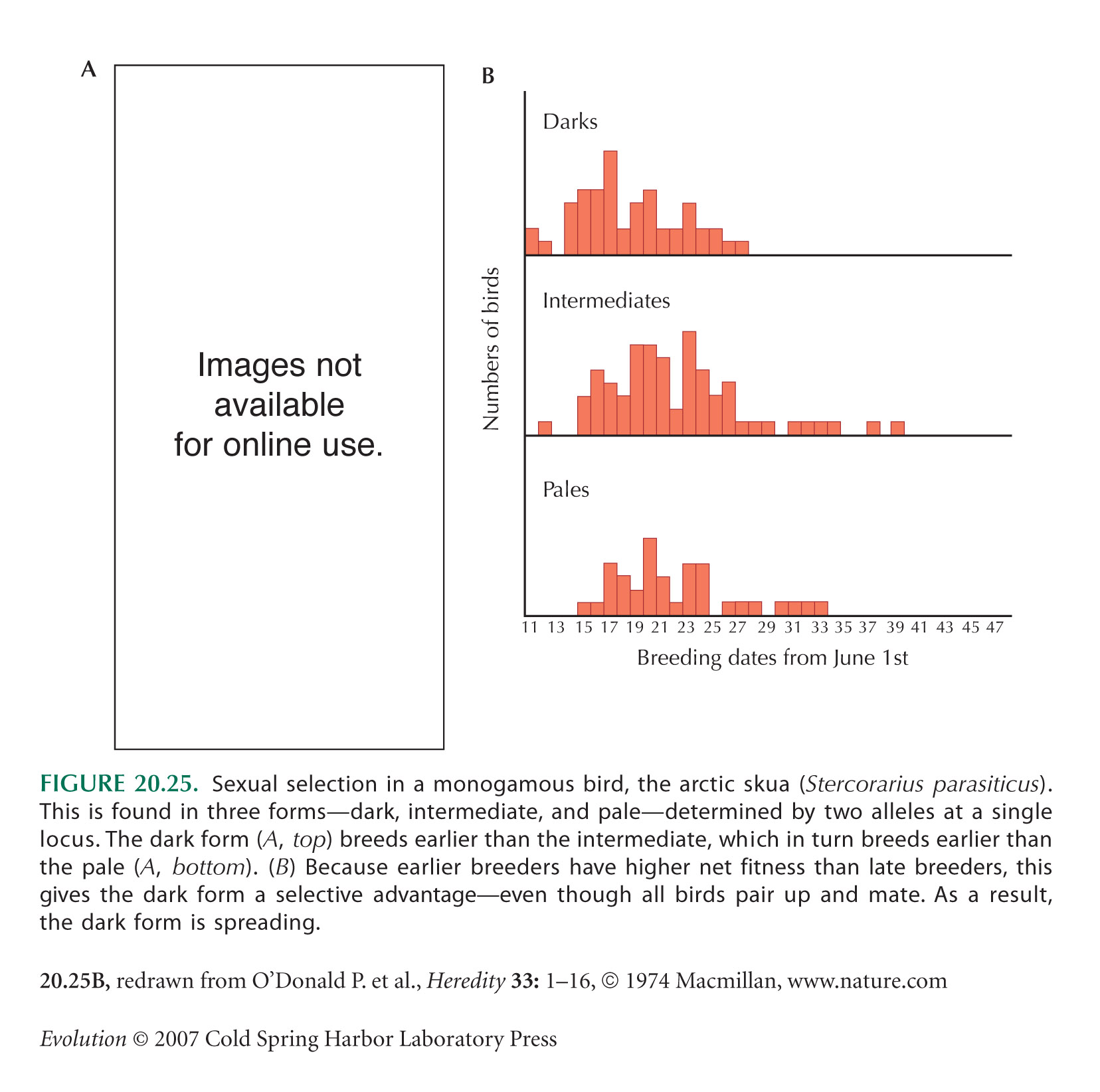
The arctic skua example of Figure 20.25 is based on a series of papers by O’Donald and colleagues (see O’Donald et al. 1974; O’Donald 1980).
Sexual Selection Involves Competition between Males and Choice of Males by Females
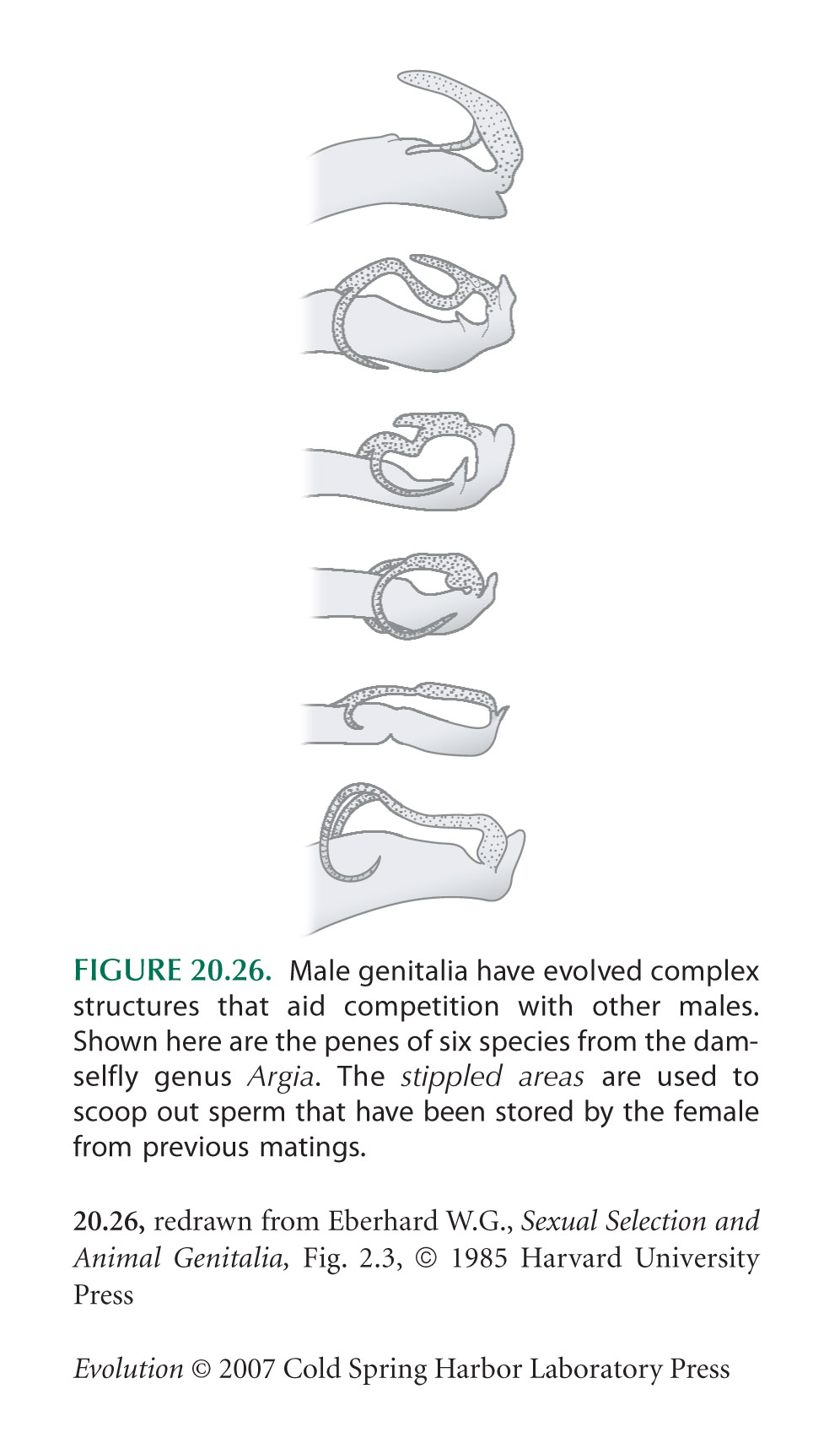
The damselfly example of Figure 20.26 is from Eberhard (1986).
The interaction between male and female genotypes seen in Figure 20.28 can maintain genetic variation (see Clark et al. 1999; Clark 2002).
Female Preferences May Evolve by Direct Selection on Females or as a Side Effect of Selection on Other Traits
Reviews of the evolution of female preference include Kirkpatrick and Ryan (1991), Andersson (1994), Ryan (1998), and Maynard Smith and Harper (2003).
The example of direct selection in blue tits comes from Norris (1990). Examples of increased fecundity due to nuptial gifts are from Butlin et al. (1987).
The nursery web spider example of Figure 20.29 is from Stalhandske (2002).
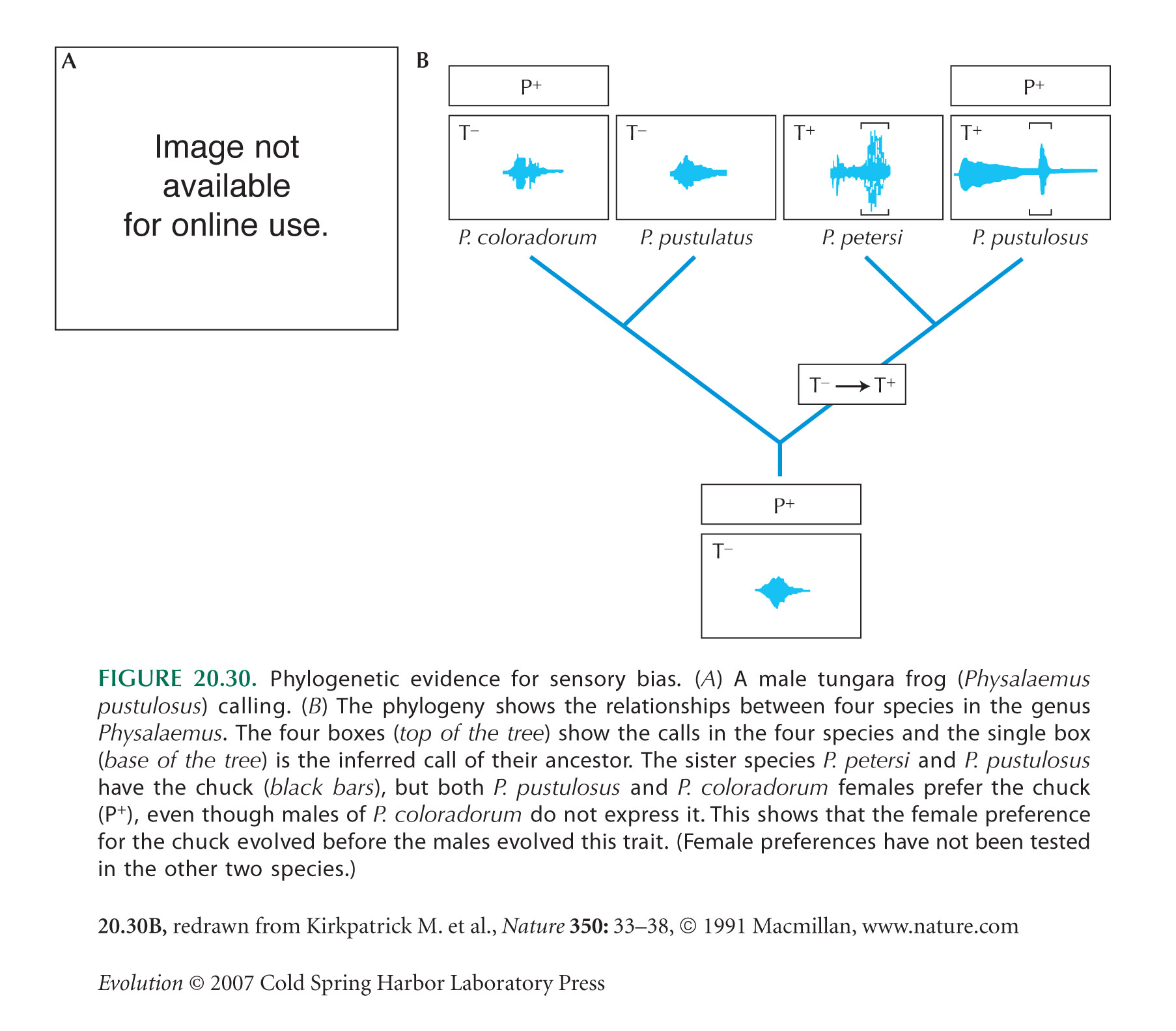
The Physalaemus example of Figure 20.30 is described by Ryan et al. (1990) and Ryan (1998). Note that in tungara frogs, there is a preference for larger males; this may be directly selected, because a larger male is more likely to be able to fertilize all the female’s eggs.
Indirect Selection on Preferences Can Lead to Fisher’s Runaway Process
Fisher (1915) first described indirect selection on female preference; the runaway process is described in Fisher (1930).
Recent work on the evolution of female preferences was stimulated by Lande’s (1981, 1982) models of sexual selection in which female preference and male trait were represented by quantitative traits. Shortly after, Kirkpatrick (1982) analyzed a similar model in which preference and trait were determined by two Mendelian genes. Lande’s (1981) model is explained .
Sexual Characteristics May Evolve to Signal Genetic Quality
Zahavi (1975) first described the handicap theory. Grafen (1990a,b) gives a clear analysis which shows that Zahavi’s theory is distinct from Fisher’s and does not rely on indirect selection.
The detailed argument that male traits may evolve as signals of male quality is quite delicate, because three sources of selection must be allowed for. The sons of females that mate with preferred males will be of higher genetic quality; they will also inherit the signal trait, which increases their mating success but reduces other components of fitness.
The association between signal trait and genetic quality may come about in two ways. The most straightforward is a so-called condition-dependent handicap. Here, males that are in good condition (partly because of heritable variation in fitness) express the signal more strongly. For example, a bird that is sick, as a result of infectious or inherited disease, may have drab plumage and be unable to make bright colors. Alternatively, there may be an epistatic handicap. Here, expression of the signal in any one male is independent of other fitness components. The association between the trait and genes that confer higher viability comes about because the survival of the more vigorous males is reduced less by the costly signal trait than is the viability of less vigorous males.
It Is Difficult to Find Evidence on Whether Females Choose Males with Good Genes
The great tit example of Figure 20.32 is from Norris (1993).
The cockroach example of Figure 20.33 is from Moore (1994).
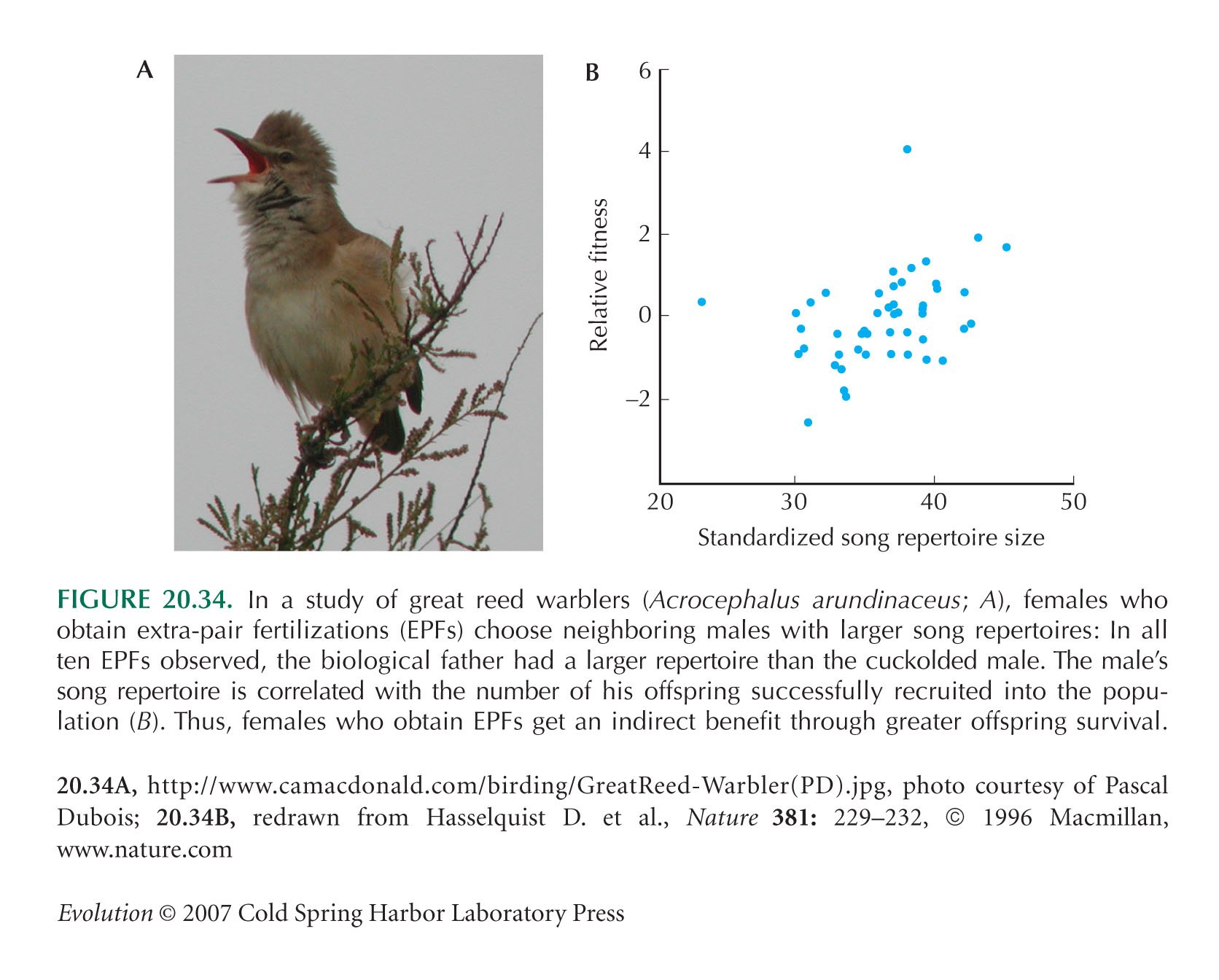
The great reed warbler example of Figure 20.34 is from Hasselquist et al. (1996).
Bro-Jørgensen et al. (2007) show that variation in swallow tail length—a classic example of a male signal (see Chapter 19 opening figure)—is due to variation in the aerodynamic optimum between individuals. Thus, it need not be a sexually selected signal of male quality.
Figures on the frequency of extra-pair fertilization are from Griffith et al. (2002).
Choice of Good Genes Requires Heritable Variation in Fitness
Hamilton and Zuk (1982) argued that coevolution between hosts and parasites is responsible for maintaining high fitness variation and that male traits evolve to signal this variation. Hamilton (1996) gives a somewhat idiosyncratic summary of the subsequent debate.
References
Andersson M. 1994. Sexual selection. Princeton University Press, Princeton, New Jersey.
Bennett A.F., Lenski R.E., and Mittler J.E. 1992. Evolutionary adaptation to temperature. I. Fitness responses of E. coli to changes in its thermal environment. Evolution 46: 16–30.
Bro-Jørgensen J., Johnstone R.A., and Evans M.R. 2007. Uninformative exaggeration of male sexual ornaments in barn swallows. Curr. Biol. 17: 850–855. [Epub 2007 Apr 5; Comment in Curr. Biol. 2007 May 15; 17: R368–R370.]
Butlin R.K., Woodhatch C.W., and Hewitt G.M. 1987. Male spermatophore investment increases female fecundity in a grasshopper. Evolution 41: 221–224.
Catchpole E.A., Morgan B.J.T., Coulson T.N., Freeman S.N., and Albon S.D. 2000. Factors influencing Soay sheep survival. J.R. Stat. Soc. Ser. C Appl. Stat. 49: 453–472.
Charlesworth B. 2000. Fisher, Medawar, Hamilton and the evolution of aging. Genetics 156: 927–931.
Clark A.G. 2002. Sperm competition and the maintenance of polymorphism. Heredity 88: 148–153.
Clark A.G., Begun D.J., and Prout T. 1999. Female × male interactions in Drosophila sperm competition. Science 283: 217–220.
Clark A.G., Aguade M., Prout T., Harshman L.G., and Langley C.H. 1995. Variation in sperm displacement and its association with accessory gland protein loci in Drosophila melanogaster. Genetics 139: 189–201.
Clutton-Brock T. 1988. Reproductive Success. University of Chicago Press, Chicago.
Cooper V.S. and Lenski R.E. 2000. The population genetics of ecological specialization in evolving Escherichia coli populations. Nature 407: 736–739.
Cooper V.S., Schneider D., Blot M., and Lenski R.E. 2001. Mechanisms causing rapid and parallel losses of ribose catabolism in evolving populations of Escherichia coli B. J. Bacteriol. 183: 2834–2841.
Darwin C.R. 1882. The descent of man, and selection in relation to sex, 2nd ed. John Murray, London.
Dawkins R. 1997. Human chauvinism. Evolution 51: 1015–1020.
Doudna J.A. and Cech T.R. 2002. The chemical repertoire of natural ribozymes. Nature 418: 222–228.
Eberhard W.G. 1986. Sexual selection and animal genitalia. Harvard University Press, Cambridge, Massachusetts.
Edwards J.S. and Palsson B.O. 2000. The Escherichia coli MG1655 in silico metabolic genotype: Its definition, characteristics, and capabilities. Proc. Natl. Acad. Sci. 97: 5528–5533.
Fisher R.A. 1915. The evolution of sexual preference. Eugen. Rev. 7: 184–192.
Fisher R.A. 1930. The genetical theory of natural selection. Oxford University Press, Oxford.
Godfray H.C.J., Partridge L., and Harvey P.H. 1991. Clutch size. Annu. Rev. Ecol. Syst. 22: 409–430.
Gould S.J. 1997. Self-help for a hedgehog stuck on a mole-hill. Evolution 51: 1020–1023.
Gould S.J. and Lewontin R. 1979. The spandrels of San Marco and the Panglossian paradigm. Proc. R. Soc. Lond. B 205: 581–595.
Grafen A. 1990a. Sexual selection unhandicapped by the Fisher process. J. Theor. Biol. 144: 473–516.
Grafen A. 1990b. Biological signals as handicaps. J. Theor. Biol. 144: 517–546.
Griffith S.C., Owens I.P.F., and Thuman K.A. 2002. Extra pair paternity in birds: A review of interspecific variation and adaptive function. Mol. Ecol. 11: 2195–2212.
Guarente L. 2001. SIR2 and aging—The exception that proves the rule. Trends Genet. 17: 391–393.
Guarente L. 2002. Ageless quest: One scientist’s search for genes that prolong youth. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Hagen D.W. and Moodie G.E.E. 1979. Polymorphism for breeding colors in Gasterosteus aculeatus. I. Their genetics and geographic distribution. Evolution 33: 641–648.
Hagen D.W., Moodie G.E.E., and Moodie P.F. 1980. Polymorphism for breeding colours in Gasterosteus aculeatus. II. Reproductive success as a result of convergence for threat display. Evolution 34: 105–1059.
Haldane J.B.S. 1941. New paths in genetics. Allen & Unwin, London.
Hamilton W.D. 1966. The moulding of senescence by natural selection. J. Theor. Biol. 12: 12–45.
Hamilton W.D. 1982. Pathogens as causes of genetic diversity. In Population biology of infectious diseases (ed. R.M. Anderson and R.M. May). Springer-Verlag, New York.
Hamilton W.D. 1996. Narrow roads of gene land, Volume 2: Evolution of sex and sexual selection. W.H. Freeman, Oxford.
Hamilton W.D. and Zuk M. 1982. Heritable true fitness and bright birds: A role for parasites? Science 218: 384–287.
Harcourt A.H., Harvey P.H., Larson S.G., and Short R.V. 1981. Testis weight, body weight and breeding system in primates. Nature 293: 55–57.
Hardin G. 1968. The tragedy of the commons. Science 162: 1243–1248.
Hasselquist D., Bensch S., and von Schantz T. 1996. Correlation between male song repertoire, extra-pair paternity and offspring survival in the great reed warbler. Nature 381: 229–232.
Hughes K.A., Alipaz J.A., Drnevich J.M., and Reynolds R.M. 2002. A test of evolutionary theories of aging. Proc. Natl. Acad. Sci. 99: 14286–14291.
Ibarra R.U., Edwards J.S., and Palsson B.O. 2002. Escherichia coli K12 undergoes adaptive evolution to acheive in silico predicted optimal growth. Nature 420: 186–190.
Jewell P.A. 1997. Survival and behaviour of castrated Soay sheep (Ovis aries) in a feral island population on Hirta, St. Kilda, Scotland. J. Zool. 243: 623–636.
Keller L. and Genoud M. 1997. Extraordinary lifespans in ants: A test of evolutionary theories of ageing. Nature 389: 958–960.
Kerr B., Riley M.A., Feldman M.W., and Bohannan B.J.M. 2002. Local dispersal promotes biodiversity in a real life game of rock–paper–scissors. Nature 418: 171–174.
Kirkpatrick M. 1982. Sexual selection and the evolution of female choice. Evolution 36: 1–12.
Kirkpatrick M. and Ryan M.J. 1991. The evolution of mating preferences and the paradox of the lek. Nature 350: 33–38.
Kirkup B.C. and Riley M.A. 2004. Antibiotic-mediated antagonism leads to a bacterial game of rock–paper–scissors in vivo. Nature 428: 412–414.
Kruuk L.E.B., Clutton Brock T.H., Slate J., Pemberton J., Brotherstone S., and Guiness F.E. 2000. Heritability of fitness in a wild mammal population. Proc. Natl. Acad. Sci. 97: 698–703.
Lande R. 1981. Models of speciation by sexual selection on polygenic traits. Proc. Natl. Acad. Sci. 78: 3721–3725.
Lande R. 1982. Rapid origin of sexual isolation and character divergence in a cline. Evolution 36: 213–223.
Lin S.J. Kaeberlin M., Andalis A.A., Sturtz L.A., Defossez P.A., Culotta V.C., Fink G.R., and Guarante L. 2002. Caloric restriction extends Saccharomyces cerevisiae lifespan by increasing respiration. Nature 418: 344–348.
Matthews D.H., Schroeder S.J., Turner D.H., and Zuker M. 2006. Predicting RNA secondary structure. In The RNA world, 3rd ed. (ed. R.F. Gesteland et al.). Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Maynard Smith J. and Harper D. 2003. Animal signals. Oxford University Press, Oxford.
Medawar P.B. 1946. Old age and natural death. Mod. Q. 2: 30–49.
Merila J. and Sheldon B. 2000. Lifetime reproductive success and heritability in nature. Am. Nat. 155: 301–310.
Moore A.J. 1994. Genetic evidence for the “good genes” process of sexual selection. Behav. Ecol. Sociobiol. 35: 235–241.
Muller H.J. 1948. Evidence on the precision of genetic adaptation. Harvey Lect. 43: 165–229.
Nee S. and Maynard Smith J. 1990. The evolutionary biology of molecular parasites. Parasitology (suppl.) 100: S5–S18.
Norris K.J. 1990. Female choice and the quality of parental care in the great tit Parus-Major. Behav. Ecol. Sociobiol. 27: 275–281.
Norris K.J. 1993. Heritable variation in a plumage indicator of viability in male great tit Parus major. Nature 362: 537–539.
O’Donald P. 1980. Genetic models of sexual selection. Cambridge University Press, Cambridge.
O’Donald P., Wedd N.S., and Davis J.W.F. 1974. Mating preferences and sexual selection in the Arctic Skua. Heredity 33: 1–16.
Paley W. 1802. Natural theology: Or, evidences of the existence and attributes of the deity, collected from the appearances of Nature. R. Fauldner, London.
Parker G.A. and Maynard Smith J. 1990. Optimality theory in evolutionary biology. Nature 348: 27–33.
Parker G.A. and Simmons L.W. 1994. Evolution of phenotypic optima and copula duration in dungflies. Nature 370: 53–56.
Partridge L. and Barton N.H. 1993. Optimality, mutation and the evolution of ageing. Nature 362: 305–311.
Partridge L. and Gems D. 2002. Mechanisms of aging: Public or private? Nat. Rev. Genet. 3: 165–175.
Piccirilli J.A., Krauch T., Moroney S.E., and Benner S.A. 1990. Enzymatic incorporation of a new base pair into DNA and RNA extends the genetic alphabet. Nature 343: 33–37.
Promislow D.E.L. 1991. Senescence in natural-populations of mammals—A comparative study. Evolution 45: 1869–1887.
Ricklefs R.E. 1998. Evolutionary theories of aging: Confirmation of a fundamental prediction, with implications for the genetic basis and evolution of life span. Am. Nat. 152: 24–44.
Roper C., Pignatelli P., and Partridge L. 1993. Evolutionary effects on age at reproduction in larval and adult Drosophila melanogaster. Evolution 47: 445–455.
Ryan M.J. 1998. Sexual selection, receiver biases, and the evolution of sex differences. Science 281: 1999–2003.
Ryan M.J., Fox J.H., Wilczynski W., and Rand A.S. 1990. Sexual selection for sensory exploitation in the frog Physalaemus pustulosus. Nature 343: 66–67.
Salehi-Ashtiani K. and Szostak J.W. 2001. In vitro evolution suggests multiple origins for the hammerhead ribozyme. Nature 414: 82–84.
Sinervo B. 1996. The rock-paper-scissors game and the evolution of alternative male strategies. Nature 380: 240–243.
Sinervo B. 2001. Runaway social games, genetic cycles driven by alternative male and female strategies, and the origin of morphs. Genetica 112: 417–434.
Singer M.C., Thomas D., and Parmesan C. 1993. Rapid human-induced evolution of insect-host associations. Nature 366: 681–684.
Snoke M.S. and Promislow D. 2003. Quantitative genetic tests of recent senescence theory: Age-specific mortality and male fertility in Drosophila melanogaster. Heredity 91: 546–556.
Stalhandske P. 2002. Nuptial gifts of male spiders function as sensory traps. Proc. R. Soc. Lond. B 269: 905–908.
Stewart E.J., Madden R., Gregory P., and Taddei F. 2005. Aging and death in an organism that reproduces by morphologically symmetric division. PLoS Biol. 3: 295–300.
Szathmary E. 1991. Four letters in the genetic alphabet: A frozen evolutionary optimum? Proc. R. Soc. Lond. B 245: 91–99.
Szathmary E. 1992. What is the optimum size for the genetic alphabet ? Proc. Natl. Acad. Sci. 89: 2614–2618.
Szathmary E. 2004. Why are there four letters in the genetic alphabet? Nat. Rev. Genet. 4: 995–1001.
Turner P.E. and Chao L. 1999. Prisoner’s dilemma in an RNA virus. Nature 398: 441–443.
Tuttle M.D. and Ryan M. 1981. Bat predation and the evolution of frog vocalizations in the Neotropics. Science 214: 677–678.
West S.A. and Herre E.A. 1998. Stabilizing selection and variance in fig wasp sex ratios. Evolution 52: 475–485.
Williams G.C. 1992. Natural selection: Domains, levels and challenges. Oxford University Press, Oxford.
Zahavi A. 1975. Mate selection—A selection of a handicap. J. Theor. Biol. 53: 205–214.
|



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}