Chapter 21 Notes
Conflict and Cooperation
Introduction
The opening figure (p. 585) shows a rufous hummingbird (Selasphorus rufus) collecting nectar from a Penstemon flower and thereby transferring pollen. We discuss the evolution of such cooperative interactions between species on pages 611–614.
It is hardly possible for any individual human, or even a family group, to be truly self-sufficient: Even a farmer depends on others to make tools, to supply seed and fertilizer, and to process and distribute the crop, not to mention clothing, education, medicine, and so on.
Social Evolution
Selection Involves Conflict between Genes and Interaction between Relatives
Strictly speaking, it would be more accurate to refer to individual traits or behaviors as selfish or altruistic, rather than talking of “selfish genes” or “altruistic genes.” Genes will always tend to maximize their own fitness (defined as the number of copies they produce), whereas individuals may evolve traits or behaviors that reduce their own fitness but help propagate their neighbor’s genes. On page 602, we explain how individuals evolve as if trying to maximize their inclusive fitness. This is a measure of individual fitness that takes into account the effects of that individual on others.
Conflict between Genes
Elements That Replicate Independently of the Host’s Chromosomes May Be Genetic Parasites
Charlesworth (2002) gives a brief review of cytoplasmic male sterility (CMS), and a commentary on Stadler and Delph’s (2002) evidence on polymorphisms in restorer alleles.
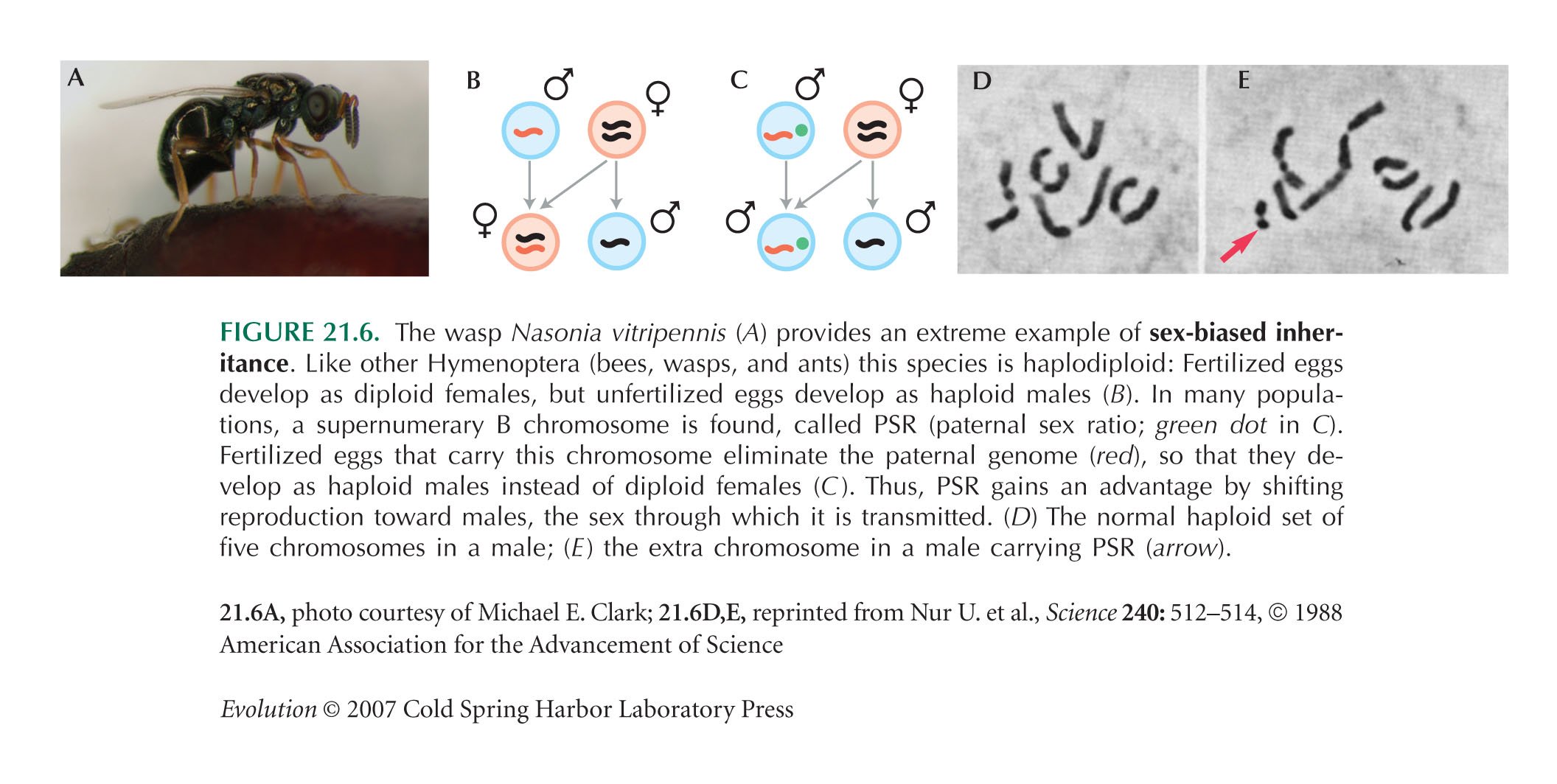
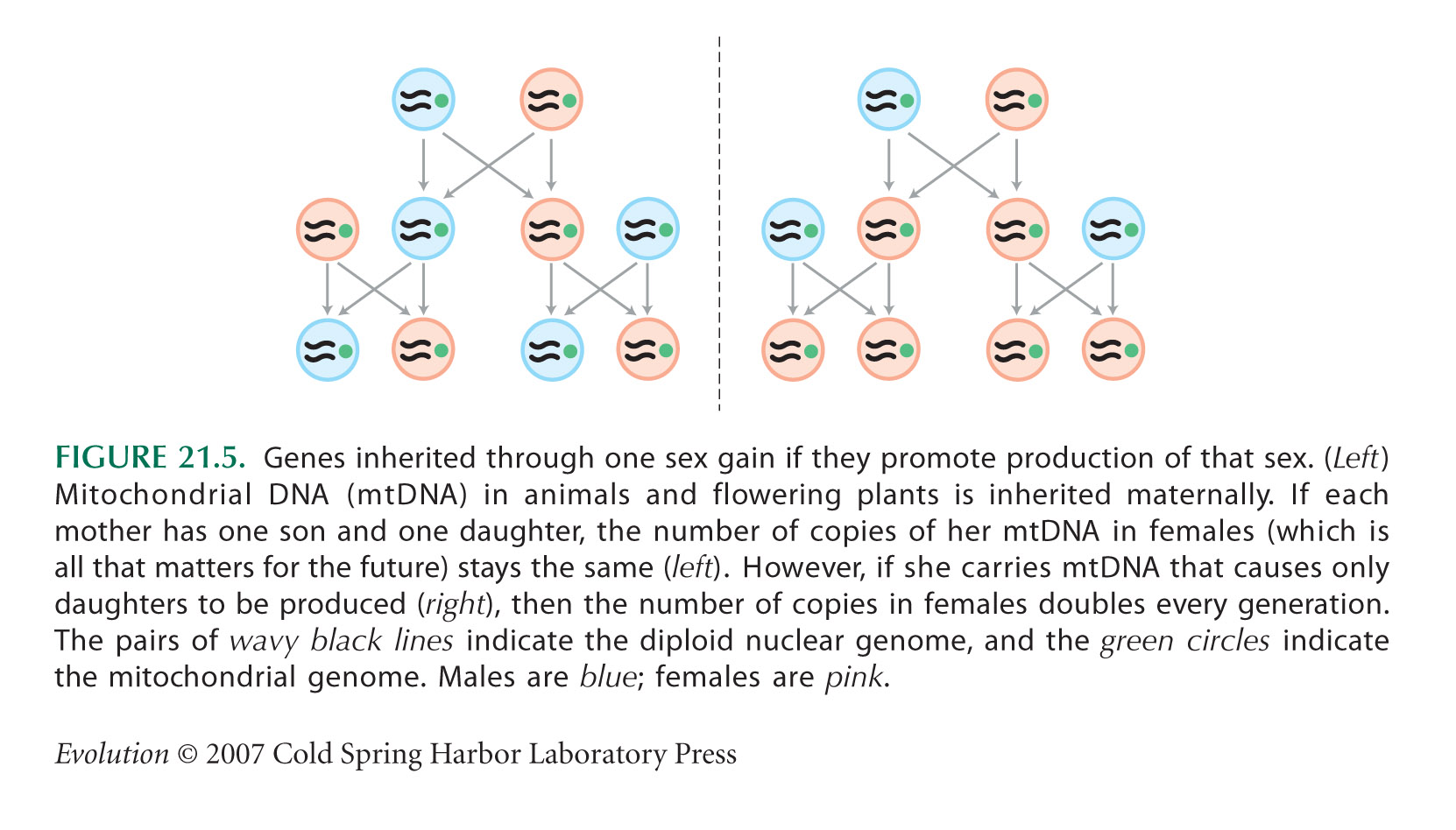
The PSR (paternal sex ratio) system in Nasonia vitripennis (Fig. 21.6) is described by Nur et al. (1988). If there is an even sex ratio, with each female producing one son and one daughter on average, as in Figure 21.5 (left), the number of copies of PSR does not increase. This is because females are then fertilizing half their eggs, so that each selfish chromosome is passed on to one offspring per female, on average. However, if the sex ratio is female biased, males have more progeny on average than females, and so PSR increases by ensuring its transmission through sons.
There Is No Clear-Cut Distinction between Genetic and Conventional Parasites
There is now a large literature on Wolbachia. Weeks et al. (2002) give a brief review, emphasizing that other endosymbionts may also be involved.
The spread of Wolbachia through California is described by Turelli et al. (1992) and Turelli and Hoffmann (1995); they model the way this parasite spreads as a wave of advance (pp. 496–499) that carries the mtDNA along with it.
Nee and Maynard Smith (1990) give an excellent account of the natural history of viruses, including defective interfering viruses (DIVs). These may increase at the expense of intact virus if only because they are shorter and therefore replicate faster. See page 569, which uses game theory to understand DIVs.
Hurst and Werren (2001) give a broad view of selfish genetic elements in eukaryotes.
Selfish Elements Often Consist of Several Tightly Linked Components
Lyon (2003) reviews the t/T system in mice. There are several distorter loci (thought to be dynein genes involved in sperm flagellae) and a single responder locus (the insensitive allele carried on t haplotypes being a fusion between two adjacent genes).
There is some divergence in putatively neutral sequence between t and + haplotypes, suggesting that this system is about 1 Myr old. However, like SD, there is little divergence in neutral sequence between t haplotypes, implying that present-day haplotypes have a very recent origin. As we will see in other examples, this suggests a continual turnover of selfish elements.
Lewontin (1962) argued that selection between demes kept the frequency of t haplotypes low in the wild—an oft-quoted example of group selection. However, Ardlie and Silver (1998) report a lower frequency of t haplotypes in large persistent populations than in small demes, which is evidence against Lewontin’s argument.
Jaenike (2001) reviews examples of meiotic drive involving the sex chromosomes.
Another well-understood example of genetic conflict is in Drosophila melanogaster.
Modifiers Evolve to Suppress Genes That Cheat
Stadler and Delph (2002) report high diversity of mtDNA variants within Silene acaulis, implying a long-standing polymorphism associated with cytoplasmic male sterility (CMS). See Charlesworth (2002). van Damme et al. (2004) and Thompson et al. (1998) give other examples of polymorphic CMS-restorer systems in Plantago and Thymus, respectively.
CMS is caused by mitochondrial variants that produce chimeric proteins, as a result of fusion between different genes. However, despite considerable research, it is not yet understood how these variants act specifically to disrupt development of pollen.
Theoretically, several outcomes are possible from the interaction between mitochondrial sterility factors and nuclear restorer alleles (Bailey et al. 2003). There may be a stable polymorphic equilibrium, continual cycling in the frequencies of cytoplasmic and nuclear variants, or a succession of substitutions of cytoplasmic sterility factors and nuclear restorers. It is not clear at present which pattern is more common.
A good example of the suppression of drive in nature involves the sex-ratio distorter in D. simulans; this is reviewed by Jaenike (2001). A driving X chromosome, XSR, can reach frequencies of up to 50%, creating a strong female-biased sex ratio. Y-linked suppressors gain an immediate advantage, because the Y competes directly with the X for transmission to sperm. Autosomal suppressors also gain an advantage if the sex ratio has become female-biased, because sons then have higher fitness than daughters (p. 506). Montchamp-Moreau et al. (2001) give evidence for the evolution of such suppressors.
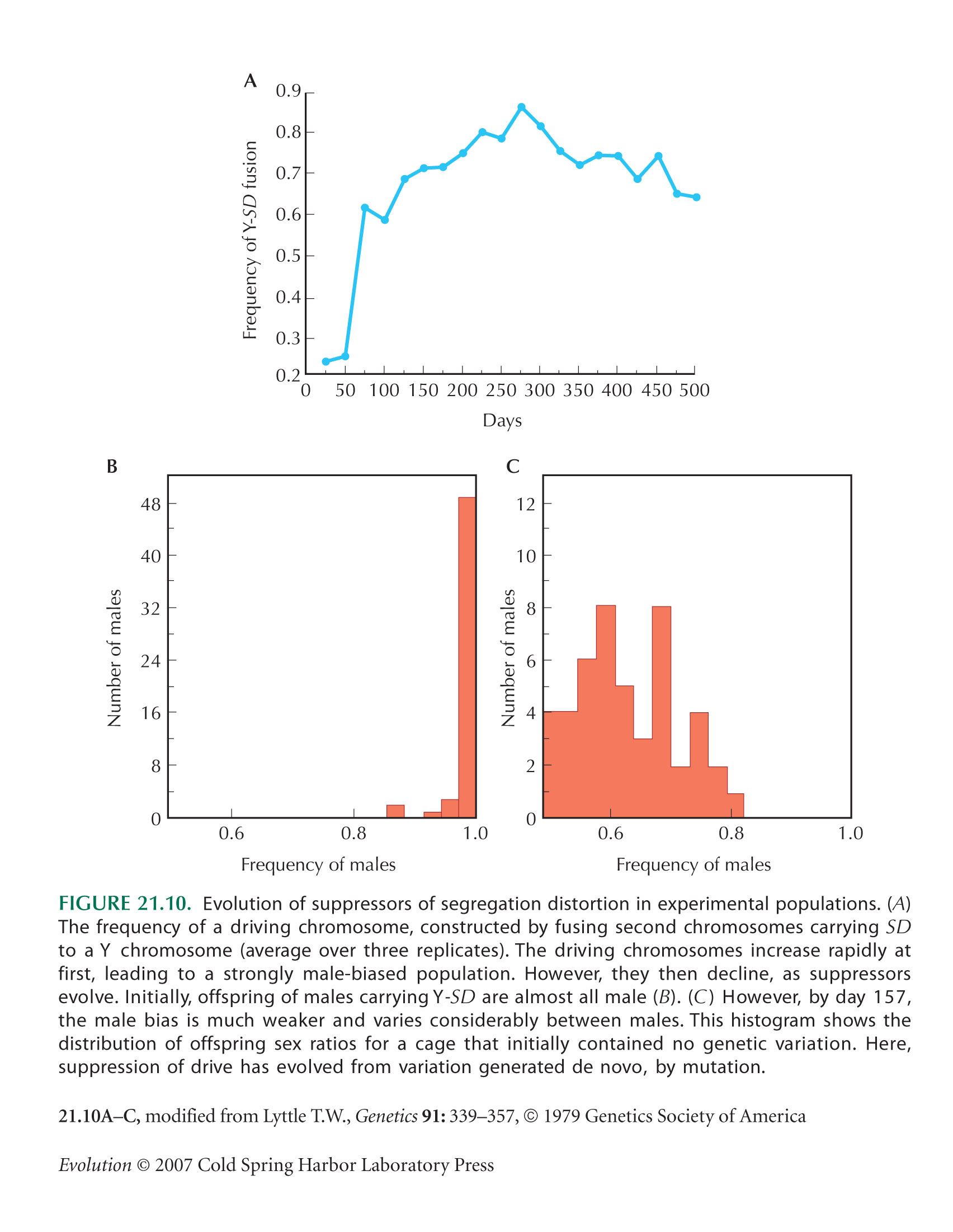
Lyttle (1979) showed the experimental evolution of suppressors of SD in laboratory populations (Fig. 21.10).
The suppression of meiotic drive in stalk-eyed flies, via sexual selection on male eyespan (p. 593), is from Wilkinson et al. (1998); see also commentary by Hurst and Pomiankowski (1998).
Repeat-induced silencing in fungi is reviewed by Selker (2002). Plasterk (2002) reviews silencing of double-stranded RNA.
When Sex Is Rare, All Genes Share the Same Interests
If a selfish element fixes throughout the host population, then it is no longer under selection to maintain its function. As we see on pages 595–597, much of the eukaryotic genome may consist of such relics. However, function can be maintained by very rare horizontal transmission between species. For example, homing endonuclease genes (HEGs) in yeast spread by inducing double-stranded breaks at specific 31-bp sites (Gimble and Thorner 1992). These breaks are repaired by copying from the HEG itself, causing a gene conversion event that increases their copy number (see Koufopanou et al. 2002 for more details). A homing endonuclease gene known as VDE is fixed within Saccharomyces cerevisae. However, across yeast species, its phylogeny does not correspond with that of the rest of the genome, showing that there have been very occasional cross-species transfers (roughly every 6 Myr; see Koufopanou et al. 2002). Such events, although rare, may be sufficient to maintain HEGs in a functional state.
The details of prokaryotic transposon and insertion copy-number regulation are from Watson et al. (2008, Chapter 11).
Evidence for the association between outbreeding and B chromosomes (Table 21.1) is from Table 1 of Burt and Trivers (1998).
Welch and Meselson (2000) set out evidence that bdelloid rotifers have reproduced asexually for extremely long times, and also lack LINE (long interspersed nucleotide element) and gypsy-like elements. However, evidence for long-term asexuality now seems less clear-cut (see Chapter 23 Web Notes).
Transposable elements can spread rapidly through populations, often with severe consequences for their hosts. We discuss some examples .
Abundance of Selfish DNA Is Determined in Several Ways
Evidence that transposable elements in Drosophila are rare and tend to be in regions of low recombination (Fig. 21.12) is from Bartolomé et al. (2002).
Table 21.2 is after Table 1 of Eickbush and Furano (2002), who discuss the differences in transposable element distribution between flies and humans.
Charlesworth et al. (1997) review the factors that regulate transposable element numbers.
Leigh (1983) introduced the idea of a “parliament of genes.”
Eukaryote Genomes Consist Largely of “Selfish” and “Junk” DNA
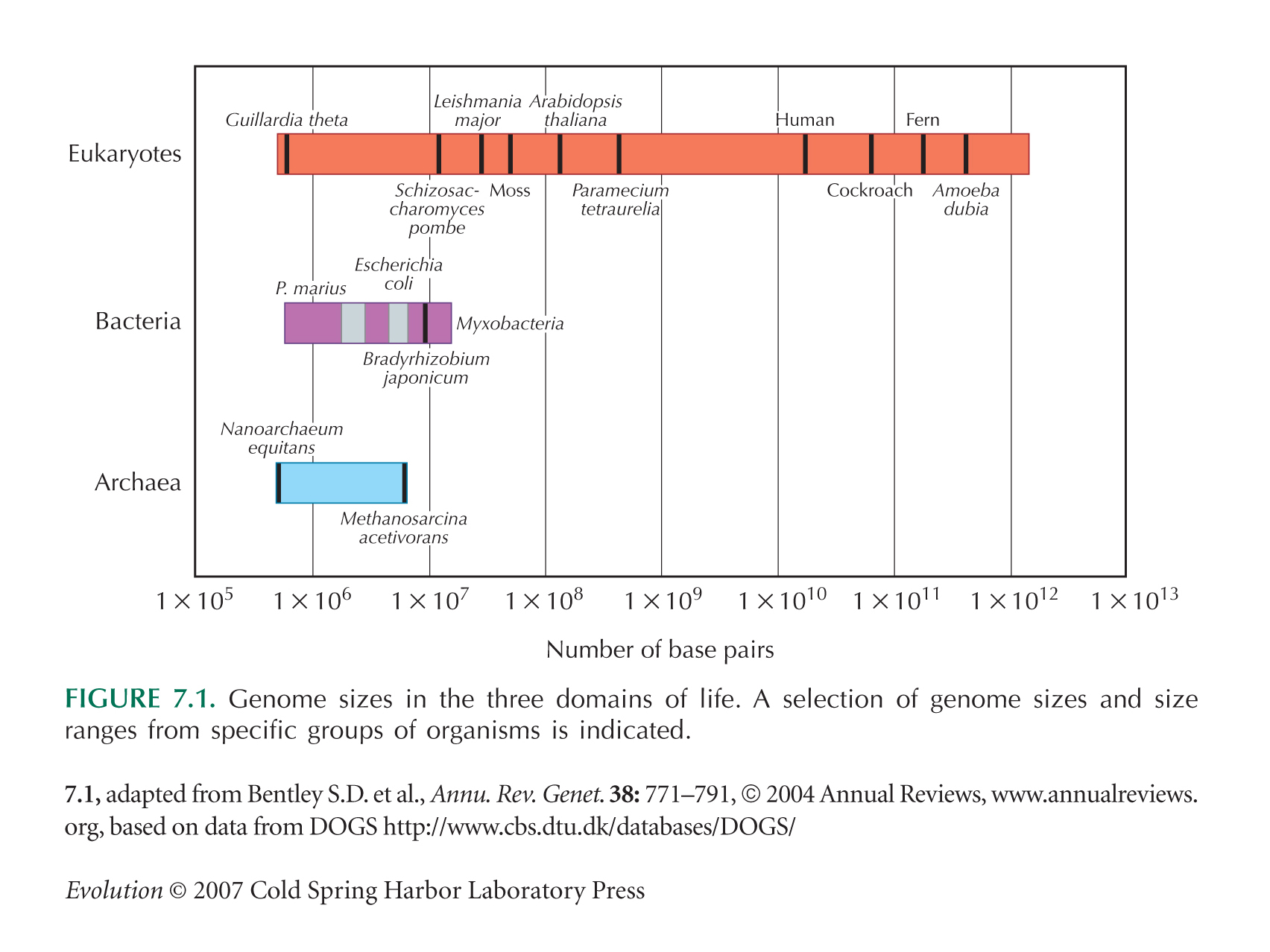
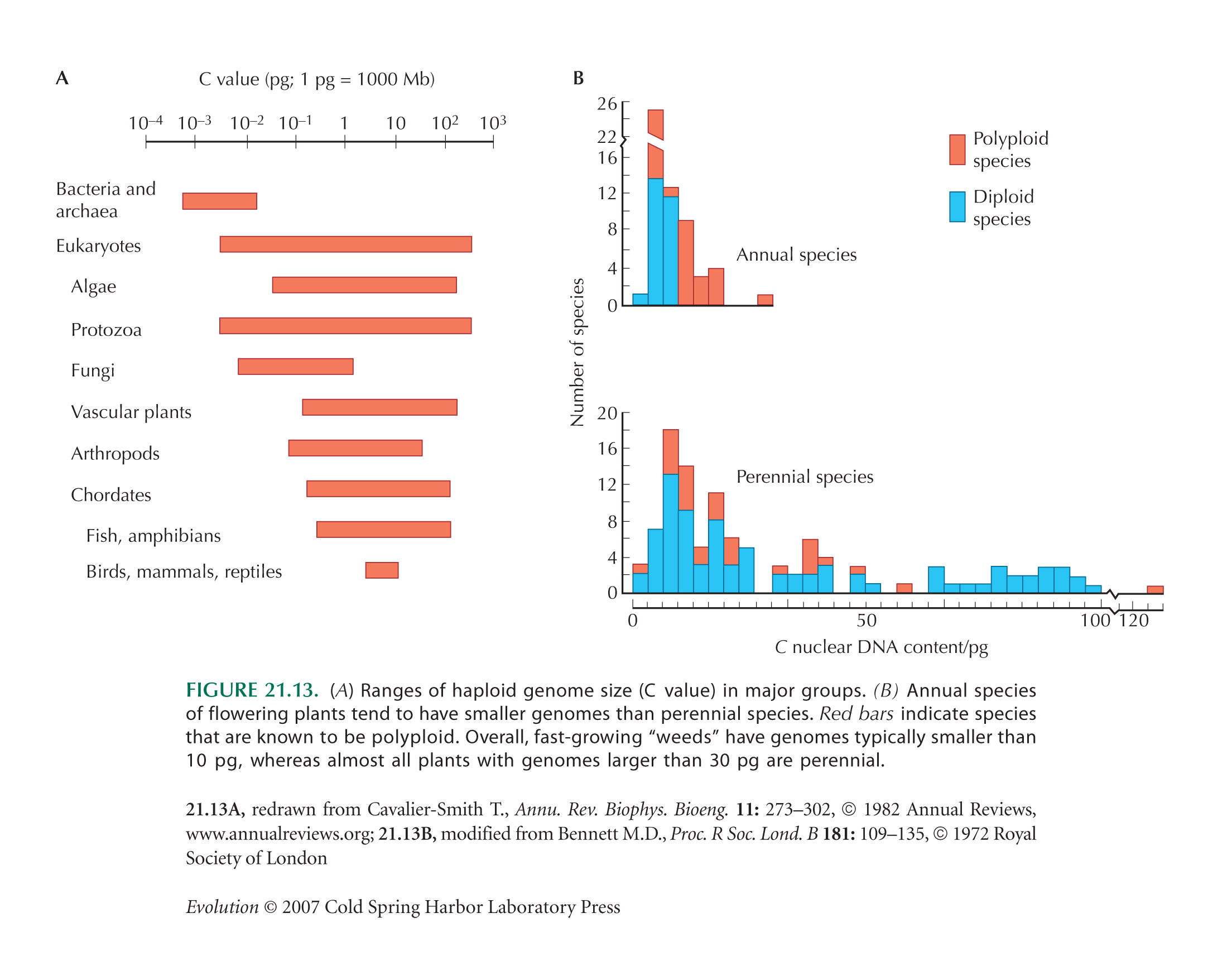
Data on genome size are from Gregory (2008). The range may be considerably greater than this: Some amoebae are reported as having genome sizes of 700 pg or more, but figures are uncertain. See Figure 7.1.
Figure 21.13A is from Figure 1 of Gregory (2001), derived from Cavalier-Smith (1982).
Variation in genome size is thought to have been important in the domestication of maize, because larger DNA content is associated with the ability to grow at lower temperatures and hence higher latitudes (Poggio et al. 1998).
Nobrega et al. (2004) made experimental deletions in mice, as cited in the text. The latter deletions had no detectable effect, even though these regions contained more than 1200 noncoding regions that are conserved between humans and rodents, and so are presumably maintained by selection (pp. 545–547).
King (2002) reviews variation in genome size in plants. Variation within maize is reported by Poggio et al. (1998) and San Miguel et al. (1998, 2000). Hatch et al. (1976) report variation in DNA content in kangaroo rats.
Greilhuber (1998) criticizes some reports of extreme genome size variation within plant species or genera, showing that they are due to technical artifacts.
Lynch and Conery (2003) include a figure that shows a smooth increase in gene number with genome size. However, this is biased by including only genomes with complete sequence, which tend to be small (see Chapter 13 Web Notes). Although the minimum genome size of a group does increase with gene number, there is no clear relationship overall.
Comparisons between annual and perennial plants (Fig. 21.13B) are from Bennett (1972).
The correlation between genome size and cell size (p. 597) is from Gregory (2001). The salamander example (Fig. 21.14) is from Roth et al. (1997); see also Bennett and Smith (1991).
Much of the Human Genome Consists of Defunct Transposable Elements
Figure 21.15 is from Figure 4 of Lynch and Conery (2003).
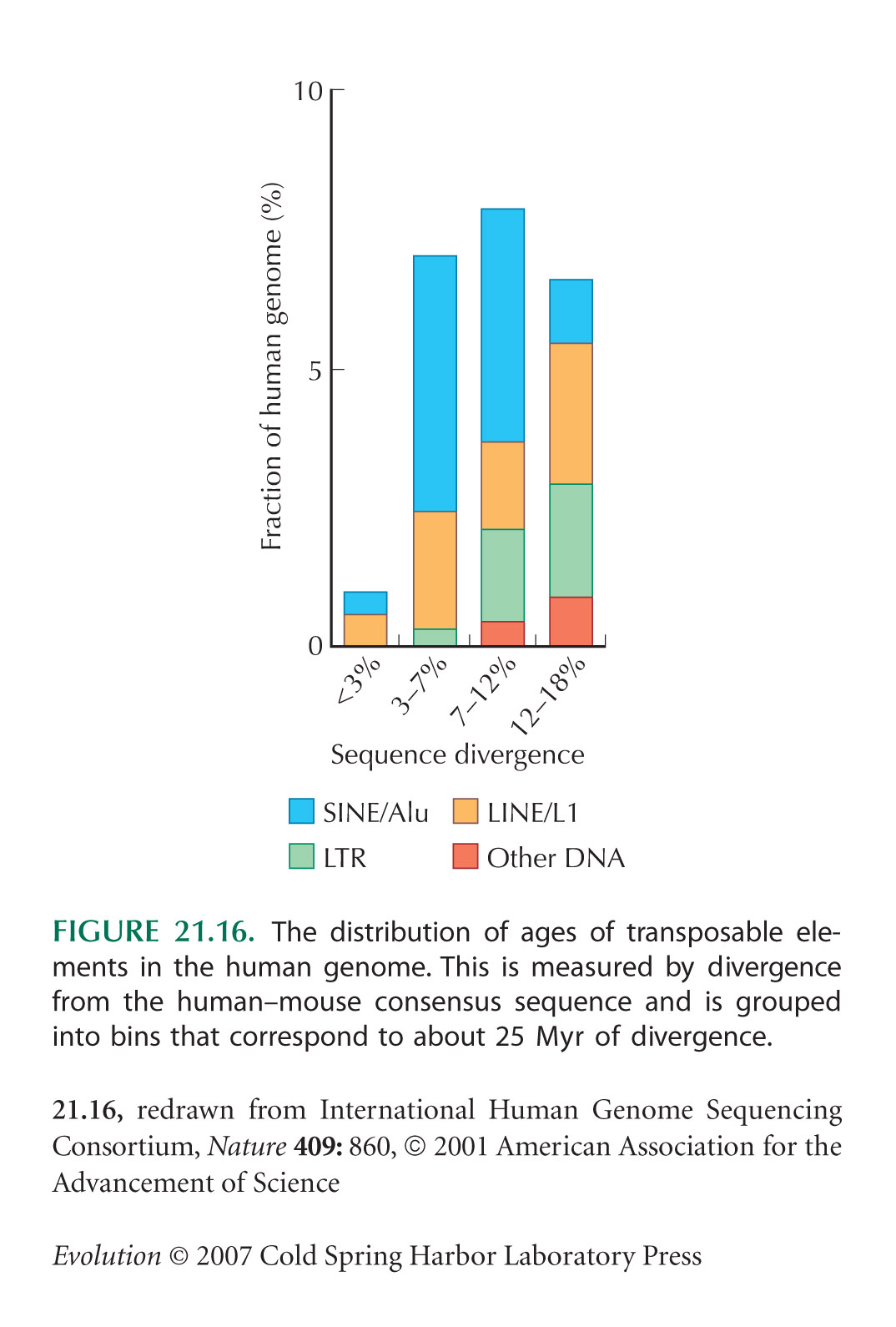
The survey of transposable elements in humans (Fig. 21.16) is from Lander et al. (2001). Only one family of LTR (long terminal repeat) retrotransposons is known to have transposed since our divergence from the chimpanzee lineage, and only a few intact copies remain. Further information on LINEs and SINEs (short interspersed nucleotide elements) can be found in Watson et al. (2008).
A substantial fraction of many eukaryotic genomes consists of simple sequence repeats (SSRs). These are tandemly repeated arrays of short sequences, from two to several hundred base pairs long (Box 13.3). We give more detail .
Transposable Elements Have Sometimes Been Co-opted to Aid Their Host
In Drosophila and other organisms, transposable elements generate much quantitative genetic variation (p. 411). This extra variation may aid host fitness. However, as argued on pages 660–662, it is highly unlikely that transposable elements could be selected in order to generate variation in this way.
In nearly every organism that has been studied, chromosome ends are capped by repeats of 5–10 bp elements—in humans, totaling 5–15 kb in length. These are maintained by telomerase, an enzyme complex that contains an RNA molecule, part of which is copied back to DNA to maintain the length of the array (pp. 214–215). Drosophila species lack this telomerase and the arrays that it maintains. Broken Drosophila chromosomes shorten by 70 bp per generation, as expected from the 8-bp length of the RNA primer on the lagging strand, and the approximately nine cell divisions in the germ line. Drosophila telomeres consist of long arrays of two classes of non-LTR retrotransposons, which maintain their length by specific transposition back onto the telomeric arrays. These retrotransposons do not insert into the interior of chromosomes, but they can attach to naked ends of DNA molecules, allowing broken chromosomes to sometimes be repaired (an illustration of this process is shown ). Intriguingly, there are similarities between the RNA molecule of telomerase and the Drosophila telomeric retrotransposons, which suggest that the latter may have evolved from telomerase itself. See Pardue and Debarshye (2003).
Interactions between Relatives
Selection Acts on the Extended Phenotype
See Dawkins (1982).
Cooperative breeding in birds is reviewed by Cockburn (1998).
Apparently Altruistic Traits Can Evolve by Kin Selection
Darwin (1859) discussed the evolution of sterile workers by what we now call kin selection in On the Origin of Species. Fisher (1930) discussed kin selection for warning color in the Genetical Theory of Natural Selection. (For recent views on the evolution of warning color, see Mallet and Joron [1999] and Servedio [2000].) Haldane (1955) outlined the idea of kin selection, but the first quantitative analyses were by Hamilton (1963, 1964a,b). Dugatkin (2007) traces this history and discusses why Fisher, Haldane, and Wright did not formulate any quantitative theory of kin selection.
We use “neighbor” loosely to refer to any individual that is influenced by the allele of interest.
Note that in the caption to Figure 21.19, we refer to the number of neighbors that carry the allele. Since these are heterozygotes, this is just “twice the allele frequency,” as stated in the text.
Several measures of relatedness can be found, including (1) probability of identity by descent (Malécot); (2) correlation between genes (Wright); and (3) regression coefficient (Hamilton). These are explained by Frank (1998) and Grafen (1985).
There is a fundamental distinction between measures of the genealogical ancestry of a gene (identical by descent or IBD, for example), and measures of the similarity of allelic state (such as the regression coefficient that appears in Hamilton’s Rule, which is analogous to linkage disequilibrium). Crozier (1987) suggests the convention that the former be called “relationship” and the latter “relatedness.” However, actual usage varies.
Sinervo and Clobert (2003) discuss the importance of kin selection in the lizard behavioral polymorphism, which we saw in Figure 20.18.
For a derivation of Hamilton’s Rule, see . This derivation can be made clearer and more general by using Price’s Equation. This is a generalization of Fisher’s Fundamental Theorem (p. 462), which partitions the effects of selection into components due to selection at different levels, and due to transmission bias. See Price (1970) and Frank (1997, 1998).
A film, WΔZ, which is loosely based on Price’s equation, has recently been released.
Inclusive Fitness Describes the Effects of Kin Selection
Hamilton’s original (1963) definition of inclusive fitness was as follows:
Inclusive fitness may be imagined as the personal fitness which an individual actually expresses in its production of adult offspring as it becomes after it has been first stripped and then augmented in a certain way. It is stripped of all components which can be considered as due to the individual’s social environment, leaving the fitness which he would express if not exposed to any of the harms or benefits of that environment. This quantity is then augmented by certain fractions of the quantities of harm and benefit which the individual himself causes to the fitness of his neighbours. The fractions in question are simply the coefficients of relationship appropriate to the neighbours whom he affects: unity for clonal individuals, one-half for sibs, one-quarter for half-sibs, one eighth for cousins...”
Grafen (1982) pointed out that most textbooks gave the wrong definition of this key quantity; he gives a clear explanation of the subtleties of the concept. Grafen (1979) shows how relatedness modifies the ESS (evolutionarily stable strategy) in the hawk–dove game (Box 20.4, p. 568).
Sex Ratio in Fig Wasps Is Determined by Both Competition and Relatedness
Some fig species are pollinated by multiple species, but this seems to be the exception. For the general biology of figs and fig wasps, see Cook and Rasplus (2003).
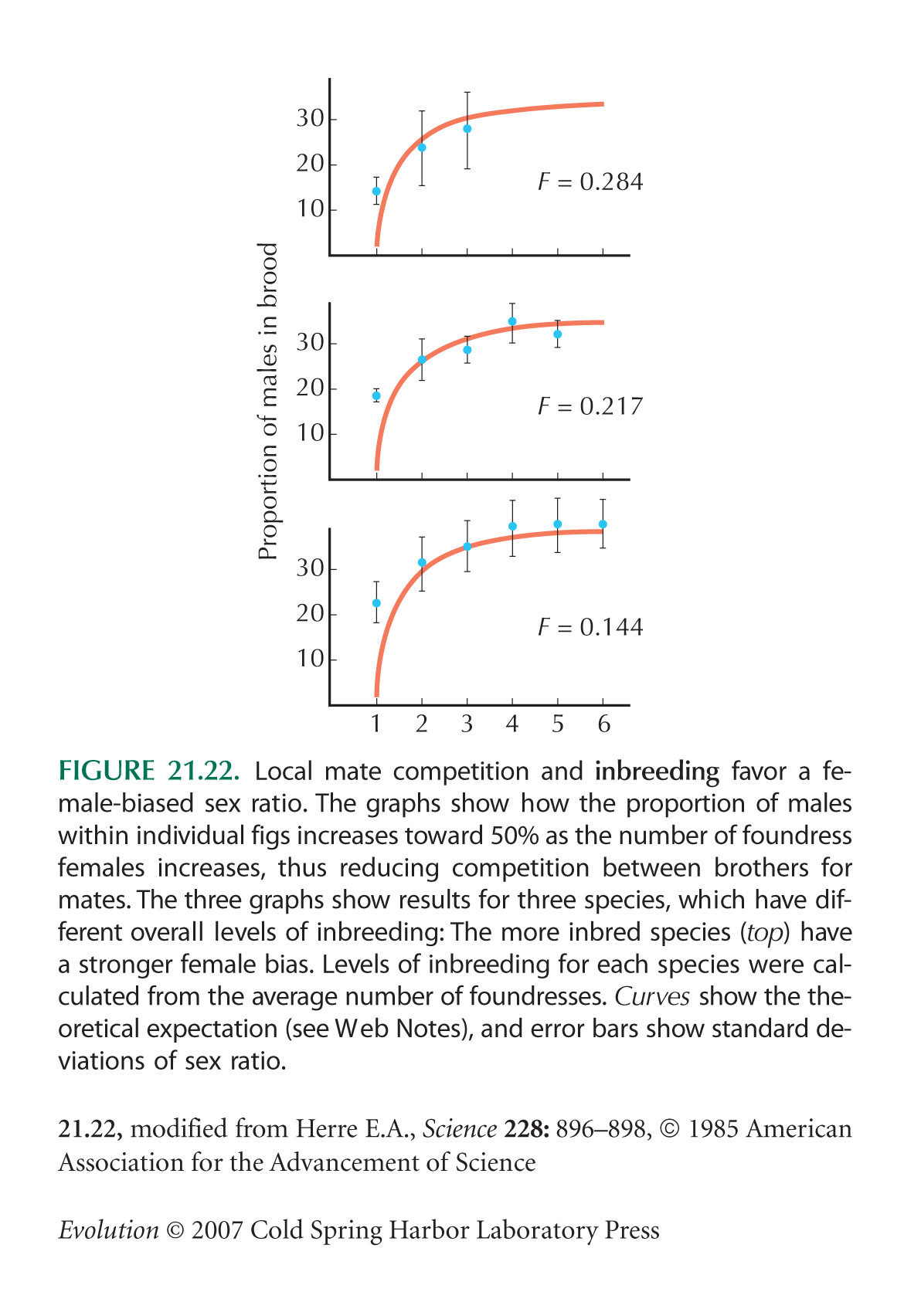
The relationship between local mate competition and sex ratio (Fig. 21.22) is from Herre (1985).
Another example of where both competition and relatedness influence behavior is the intensity of aggression between male . In species that have few foundresses, there is strong intermale competition as well as high relatedness. These opposing effects cancel, and so there is no correlation between the relatedness among males and the level of aggression (see ). However, there is a strong negative correlation with the average number of females that develop within the fruit: If there are only a few females available, then it is worth fighting for them (see ). (The level of aggression was measured by scoring injuries to the males who remain after females have left the fruit. For example, loss of an antenna counts as 1/2, whereas loss of a head and evisceration count as 8.)
To see how the equilibrium sex ratio can be predicted, as in Figure 21.22, see Problem 21.7.
Griffin and West (2002) explain that competition between relatives must be taken into account, along with kin selection.
Parents and Offspring Have Different Evolutionary Interests
Parker and MacNair (1978a,b) set out the theory of parent–offspring conflict.
Godfray et al. (1991) review experiments on clutch size manipulation.
Recall that in the previous chapter (pp. 575–578), we discussed the conflict between males and females that arises from sexual selection. Partridge and Hurst (1998) discuss sexual selection from this point of view.
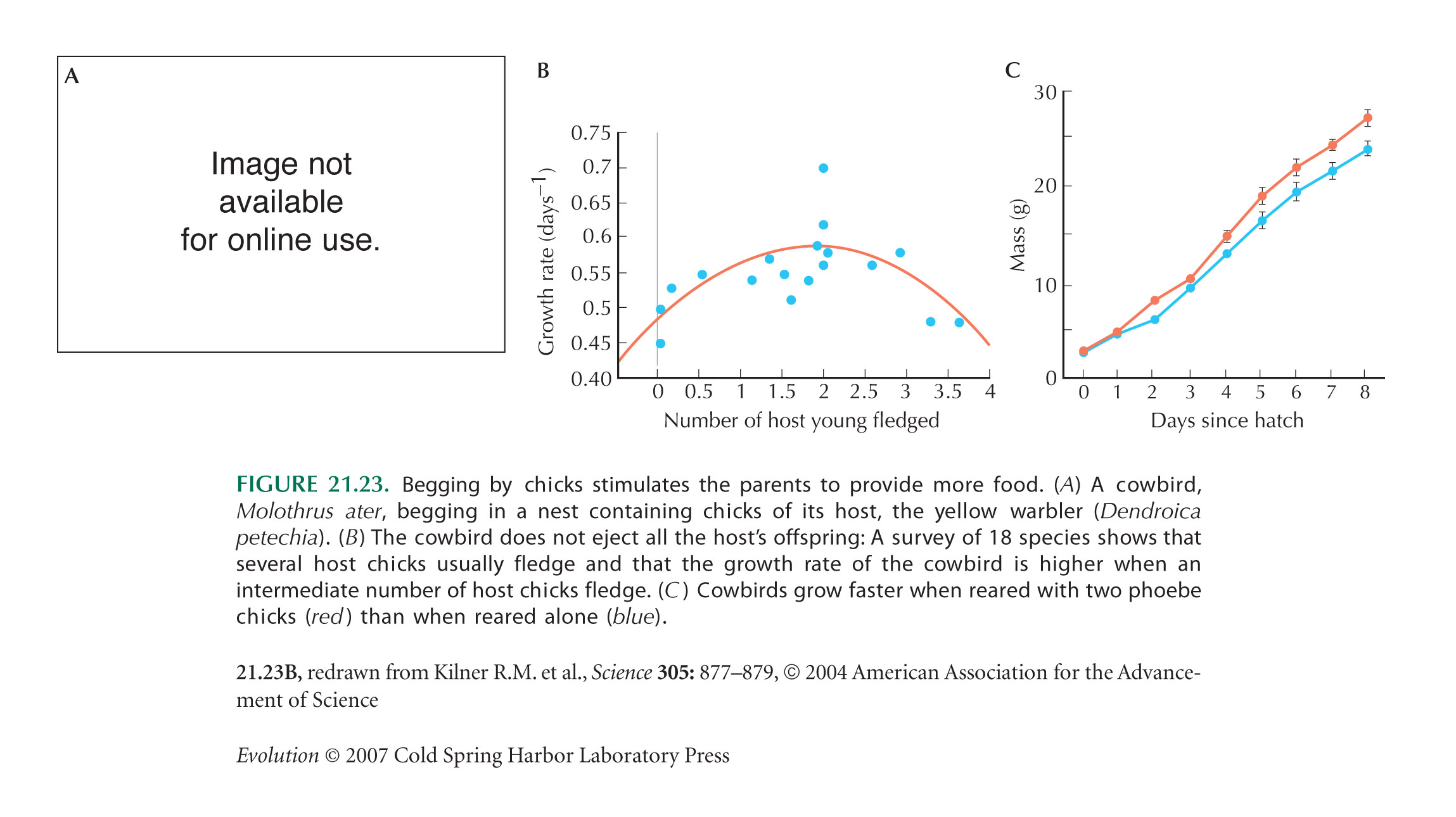
The example of begging by chicks (Fig. 21.23) is from Kilner et al. (2004).
Spencer et al. (1999), Spencer (2000), Sleutels and Barlow (2002), and de la Casa-Esperón and Sapienza (2003) review genomic imprinting. Smith and Hurst (1998) compare rates of molecular evolution in imprinted and nonimprinted genes.
We refer to genomic imprinting to distinguish this phenomenon from the behavioral imprinting of offspring on their parents.
For details of the molecular mechanism of imprinting(Fig. 21.24), see Watson et al. (2008). A regulatory sequence that silences the gene is methylated in sperm but not in eggs. Methylation stops the silencing effect, allowing expression of the paternal allele. This pattern is preserved through subsequent cell divisions.
Sometimes, Relatives Can Be Recognized
The long-tailed tit example is from Russell and Hatchwell (2001).
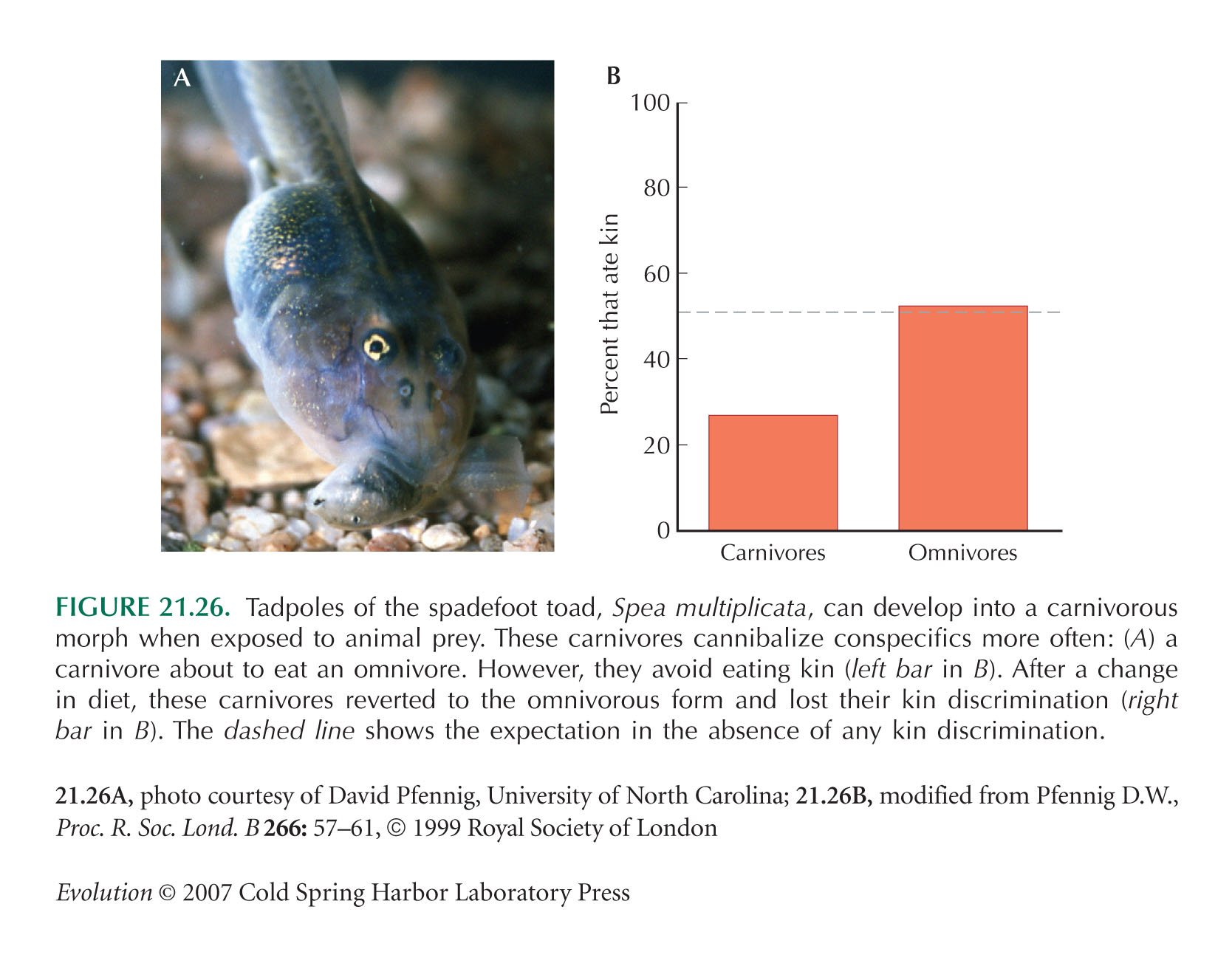
The spadefoot toad example (Fig. 21.26) is from Pfennig (1999).
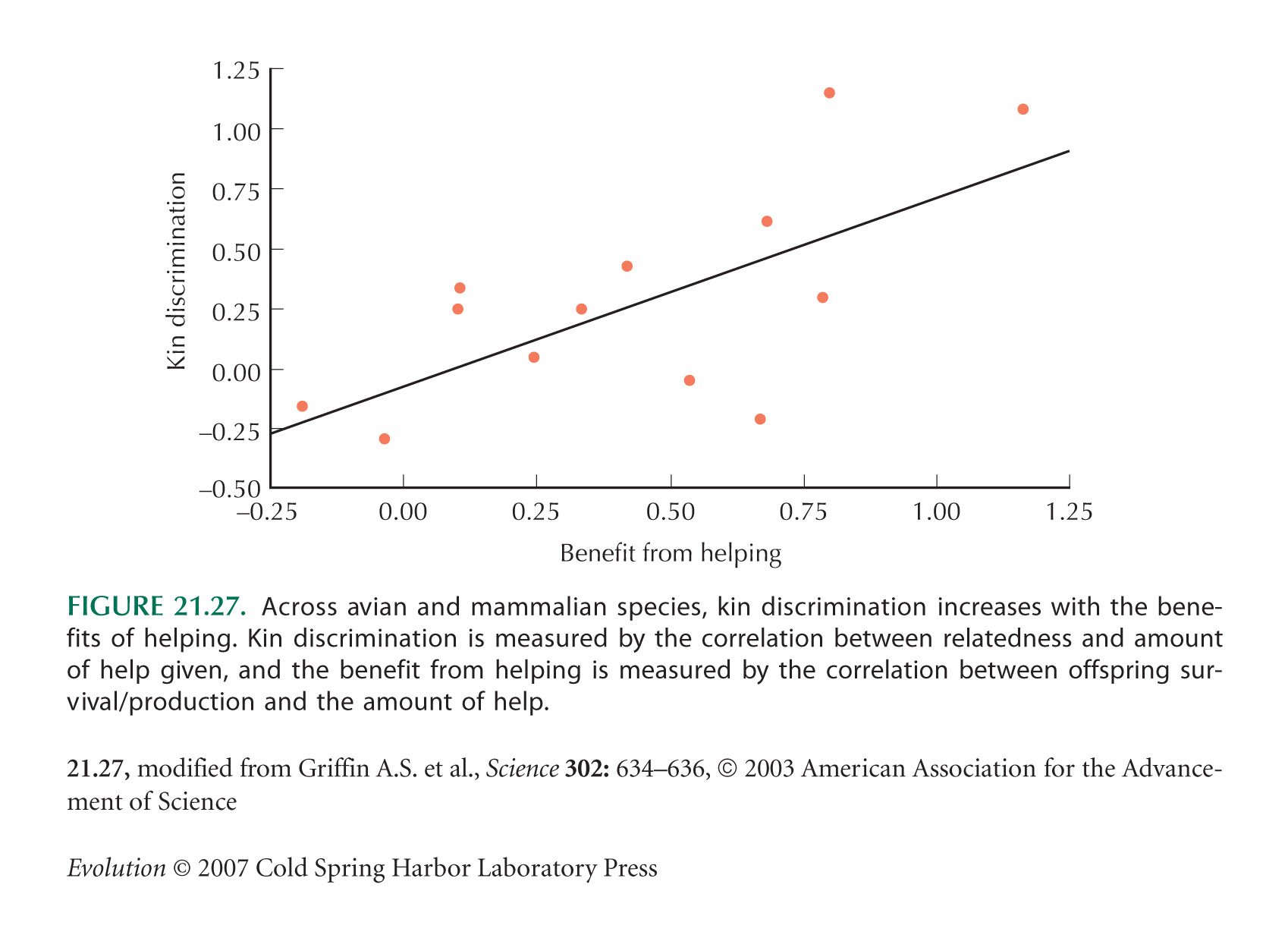
The survey of kin discrimination (Fig. 21.27) is from Griffin and West (2003).
Brookfield (1998) reviews some examples of quorum sensing, including the potato pathogen example. Diggle et al. (2007) show how cooperative quorum sensing might be exploited and suggest that it could be stabilized by kin selection; see also Sandoz et al. (2007).
Kin selection arises when interacting individuals share alleles. The most obvious reason why neighbors should share alleles is that they share ancestry. We discuss other reasons .
Selection among Groups Is Weaker than Selection among Individuals
As explained on page 607, the weakness of group selection, relative to selection among individuals, was emphasized in the 1960s; in parallel, the closely related concept of kin selection was developed. Reviews of group selection from this period include Leigh (1983), Maynard Smith (1976), Williams (1992), and Wilson (1983). Recently, Wilson and Wilson (2007a,b) have revived arguments over the relative roles of kin and group selection and the distinction (if any) between them. For alternative viewpoints, see Lehmann et al. (2007) and the exchange between West et al. (2007, 2008) and Wilson (2008).
Wright’s Shifting Balance Involves Selection among Alternative Adaptive Peaks
Wright (1931, 1932) first set out the shifting balance theory (SBT) and reiterated it in many subsequent papers (see Provine 1986a). Provine (1986b) gives an excellent commentary, and Coyne et al. (1997, 2000) provide critical reviews.
There is a particularly close analogy between simulated annealing (Kirkpatrick et al. 1983) and the SBT.
Another difficulty facing the SBT is that there is little or no recombination between alternative adaptive peaks. These alternatives will differ in many respects (as we see in nature, where hybrid zones separate highly divergent populations). Thus, selection can only choose between a small number of types, each differing by many characteristics. Selection between sexually reproducing individuals can be much more efficient, because it can separate out the specific genetic loci that actually increase fitness. We explain why recombination makes selection more efficient in Chapter 23.
Wright used the desert annual plant Linanthus parryae as an example of his “shifting balance”: He thought that variation in the frequency of flower color polymorphism from place to place was due to random drift. However, Schemske and Bierzychudek (2001, 2007) show that this geographic variation is in fact due to selection. See commentary by Pennisi (2007).
Selection Can Act among Species but Is Exceedingly Slow
Gould and Eldredge (1977) proposed a theory of punctuated equilibrium, in which morphological change occurs at brief speciation events, which punctuate long periods of stasis. Under this theory, long-term trends are due to species selection as well as to biases in the direction of change at speciation. See also Stanley (1975, 1979).
Coyne and Orr (2004) summarize comparative evidence on factors affecting speciation rates. The study cited in Figure 21.29 is from Arnqvist et al. (2000).
Table 21.3 is from Maynard Smith and Szathmary (1995).
Evolution of Cooperation
Cooperation May Be to the Participants’ Mutual Advantage
The examples in this section are discussed in reviews by Griffin and West (2003) and by Clutton-Brock (2002).
Clutton-Brock et al. (1998) document the large weight losses by babysitting meerkats.
Mutualisms might benefit both partners, but often there is the possibility of taking from the partner without giving in return, as in the Prisoner’s Dilemma. Simply because the two partners mutually benefit from cooperation does not mean that the problem of cooperation is solved.
The “honest signaling” described in the previous chapter (p. 581) is, in effect, a form of cooperation.
Grafen (1979) incorporated relatedness into the hawk–dove game.
Box 21.2 Social Insects
It is surprising that such complexity can evolve despite the genetic heterogeneity within a social colony, which makes it vulnerable to subversion from within. However, as we see from the policing of workers, this heterogeneity between workers means that there is more incentive to police, and such repression of competition is often more potent than kin-selected self-restraint. Thus, there can be more reproductive harmony in those colonies that have more heterogeneity.
Anderson and McShea (2001) review social organization in ants, and Oster and Wilson (1979) review social insects more generally. The classic works on the social insects, in an evolutionary context, are Wilson (1971, 1975). Maschwitz and Maschwitz (1974) describe how exploding workers is a nest defense.
Competition Can Be Suppressed by Reciprocation, Punishment, and Policing
Axelrod and Hamilton (1981) tested a wide range of strategies and found that tit-for-tat (TFT) consistently won the Prisoner’s Dilemma. In fact, this strategy can be proved to be an ESS. For more background, see and Hamilton (1996a,b).
The classic comparative test of the relationship between policing and multiple mating is honey bee (Apis mellifera) versus bumble bee (Bombus terrestris). Bumbles are monandrous, have no policing, and have much worker reproduction, whereas honey bees are polyandrous, have worker policing, and less than 1/1000 surviving eggs are laid by workers. However, this is only a single comparison. For reviews of policing in social insects, see Ratnieks and Wenseleers (2005) and Ratnieks (1988).
we explain why policing is favored in species where the queen mates many times.
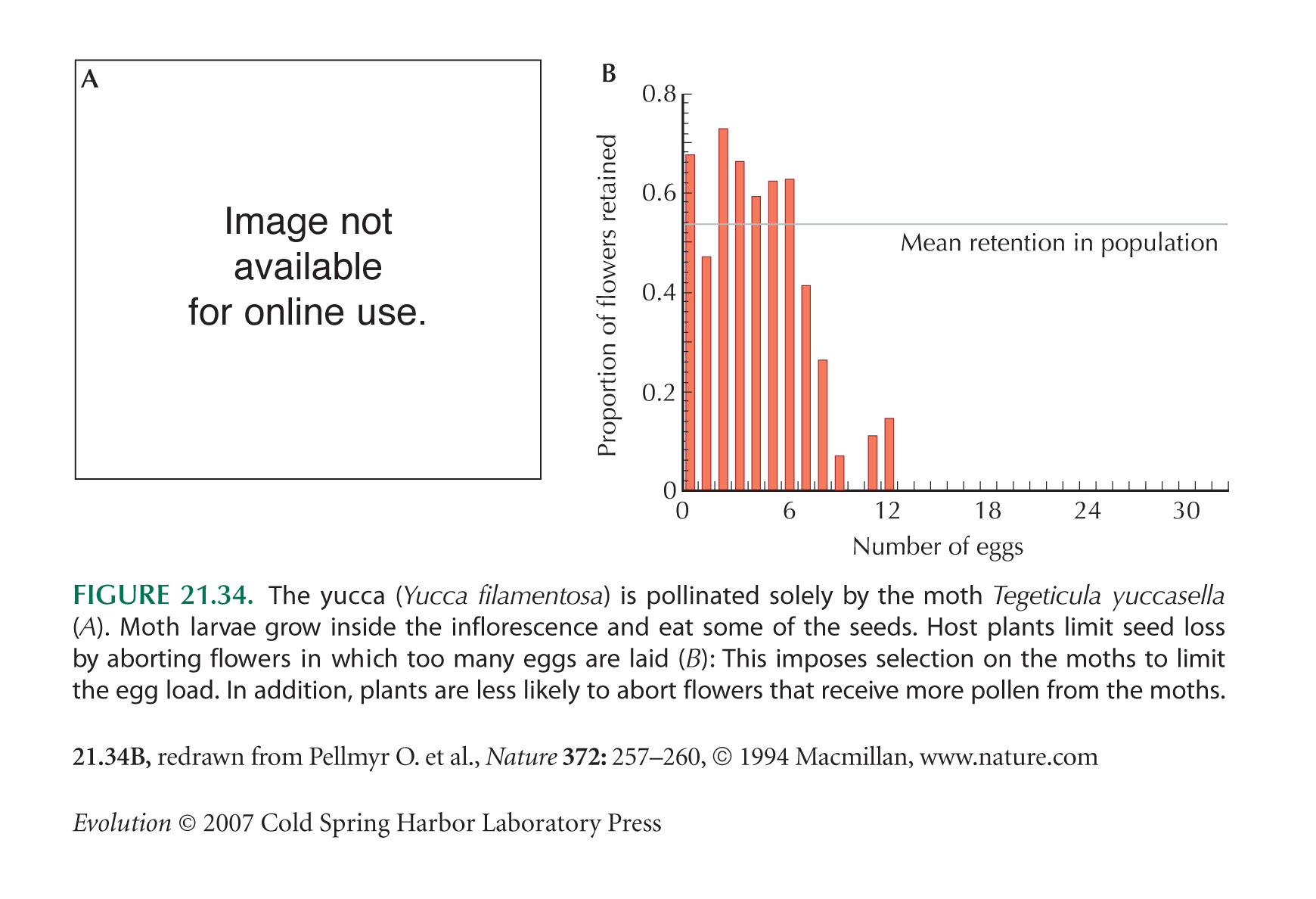
The yucca/yucca moth example (Fig. 21.34) is from Pellmyr and Huth (1994) and Pellmyr and Leebensmack (1999); the latter use the molecular clock to date mutualism at 40 Myr.
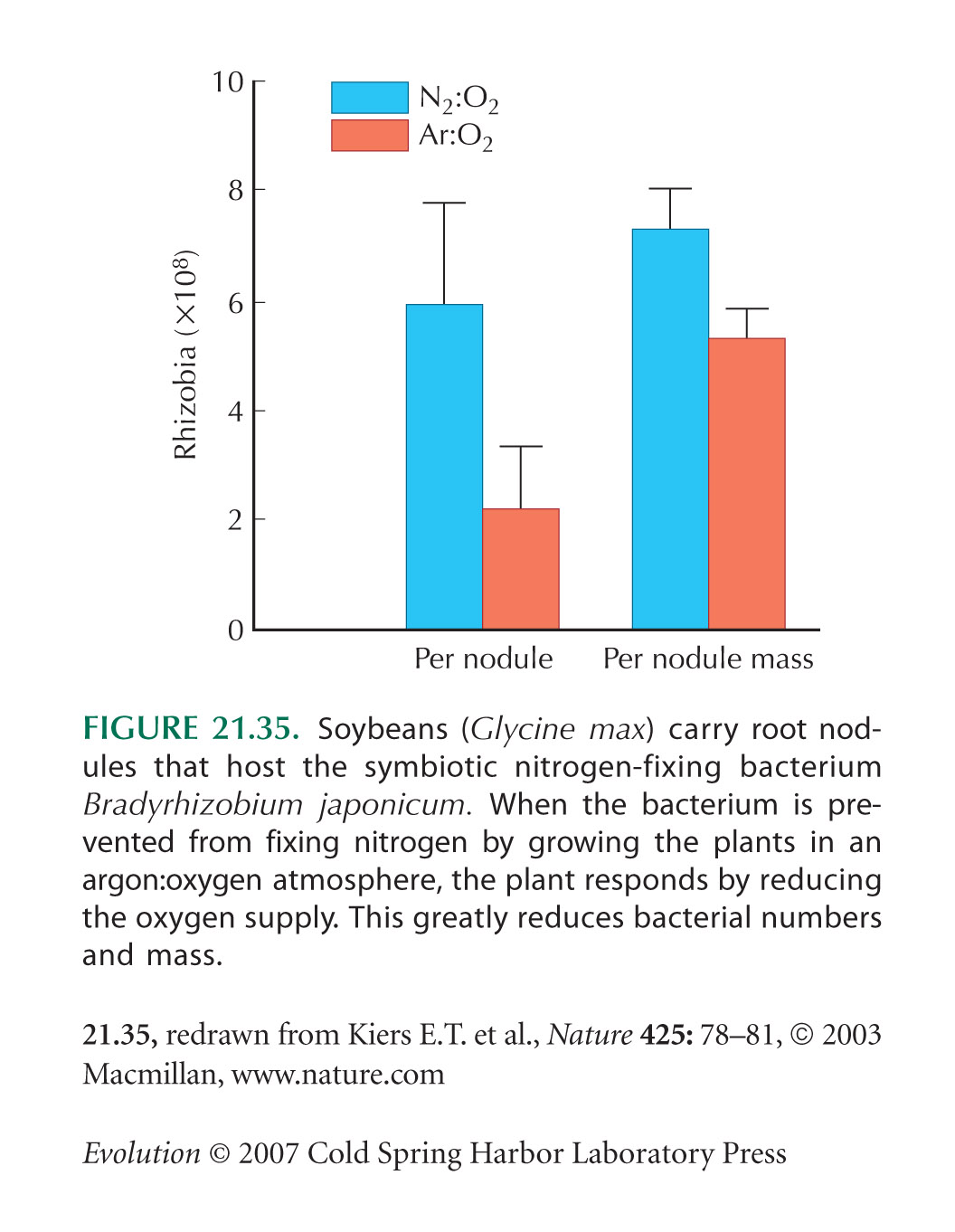
The nitrogen fixation example (Fig. 21.35) is from Kiers et al. (2003).
The colonial hydrozoan example is from Hart and Grosberg (1999).
Pfeiffer and Bonhoeffer (2003) discuss the origins of multicellularity. See also Chapters 9 and 24 and Maynard Smith and Szathmary (1995).
Conflict and Cooperation Were Important in Early Evolution
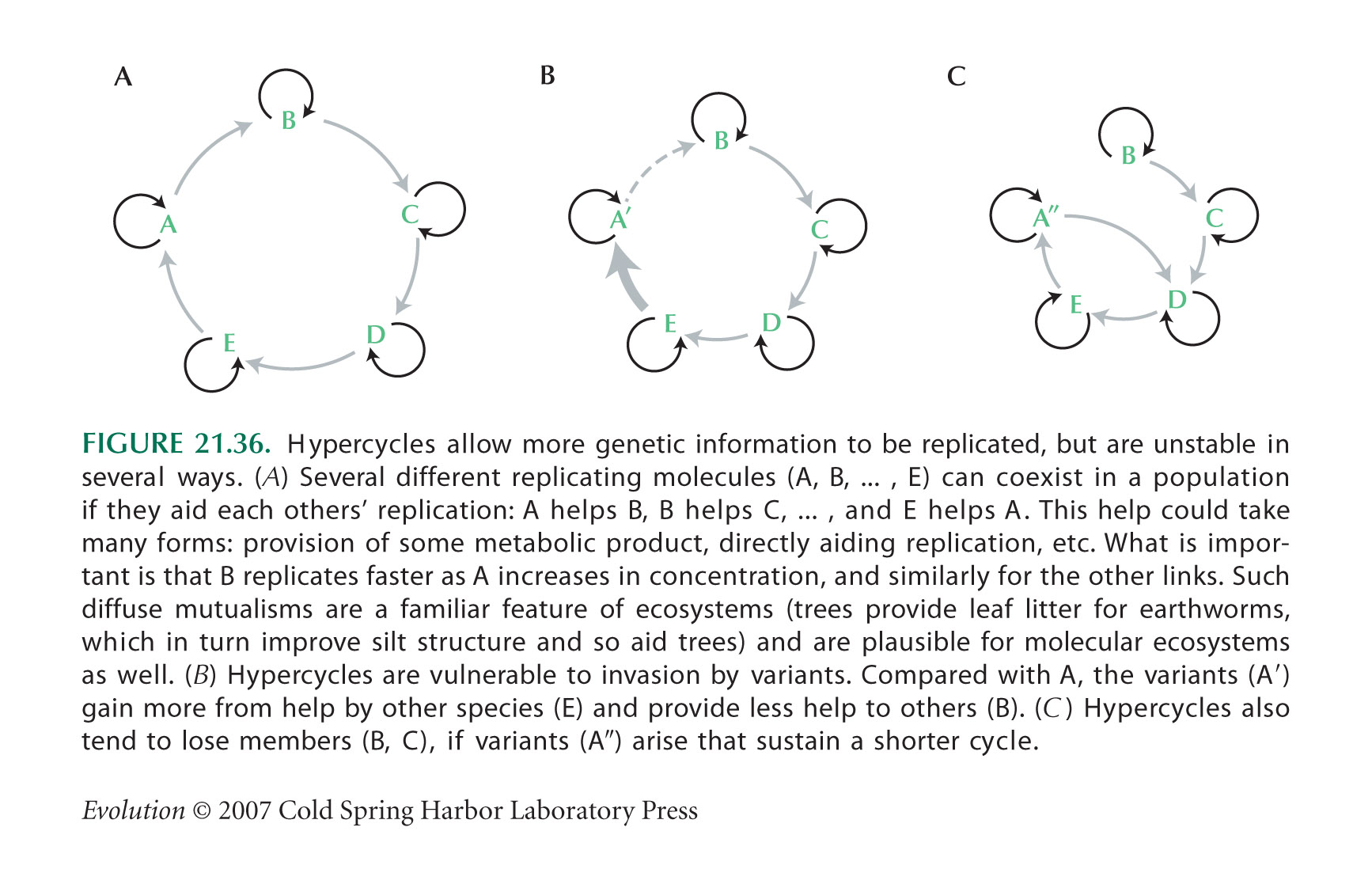
Hypercycles (Fig. 21.36) are discussed by Eigen et al. (1981) and by Eigen (1992).
The stochastic corrector model (Fig. 21.37) is from Maynard Smith and Szathmary (1995).
Summary
The argument that most natural selection is due to the struggle between host and parasite was made by Haldane (1949) and strongly promoted by W.D. Hamilton, especially as the process driving female mate choice and the evolution of sexual reproduction. See page 583 and Hamilton (1996b).
References
Anderson C. and McShea D.W. 2001. Individual versus social complexity, with particular reference to ant colonies. Biol. Rev. Camb. Philos. Soc. 76: 211–238.
Ardlie K.G. and Silver L.M. 1998. Low frequency of t haplotypes in natural populations of house mice (Mus musculus domesticus). Evolution 52: 1185–1196.
Arnqvist G., Edvardsson M., Friberg U., and Nilsson T. 2000. Sexual conflict promotes speciation in insects. Proc. Natl. Acad. Sci. 97: 10460–10464.
Axelrod R. and Hamilton W.D. 1981. The evolution of co-operation. Science 211: 1390–1396.
Bailey M.F., Delph L.F., and Lively C.A. 2003. Modeling gynodioecy: Novel scenarios for maintaining polymorphism. Am. Nat. 161: 762–776.
Bartolomé C., Maside X., and Charlesworth B. 2002. On the abundance and distribution of transposable elements in the genome of Drosophila melanogaster. Mol. Biol. Evol. 19: 926–937.
Bennett M.D. 1972. Nuclear DNA content and minimum generation time in herbaceous plants. Proc. R. Soc. Lond. B 181: 109–135.
Bennett M.D. and Smith J.B. 1991. Nuclear DNA amounts in angiosperms. Philos. Trans. R. Soc. Lond. B 334: 309–345.
Brookfield J.F.Y. 1998. Quorum sensing and group selection. Evolution 52: 1263–1269.
Burt A. and Trivers R. 1998. Selfish DNA and breeding system in flowering plants. Proc. R. Soc. Lond. B 265: 141–146.
Cavalier-Smith T. 1982. Skeletal DNA and the evolution of genome size. Annu. Rev. Biophys. Bioeng. 11: 273–302.
Charlesworth B., Langley C.H., and Sniegowski P.D. 1997. Transposable element distributions in Drosophila. Genetics 147: 1993–1995.
Charlesworth D. 2002. What maintains male-sterility factors in plant populations? Heredity 89: 408–409.
Clutton-Brock T. 2002. Breeding together: Kin selection and mutualism in cooperative vertebrates. Science 296: 69–72.
Clutton-Brock T.H., Gaynor D., Kansky R., Maccoll A.D.C., McIlrath G., et al. 1998. Costs of cooperative behaviour in suricates (Suricata suricatta). Proc. R. Soc. Lond. B 265: 185–190.
Cockburn A. 1998. Evolution of helping behaviour in cooperatively breeding birds. Annu. Rev. Ecol. Syst. 29: 141–177.
Cook J.M. and Rasplus J.Y. 2003. Mutualists with attitude: Coevolving fig wasps and figs. Trends Ecol. Evol. 18: 241–248.
Coyne J.A. and Orr H.A. 2004. Speciation. Sinauer Press, Sunderland, Massachusetts.
Coyne J.A., Barton N.H., and Turelli M. 1997. A critique of Wright’s shifting balance theory of evolution. Evolution 51: 643–671.
Coyne J.A., Barton N.H., and Turelli M. 2000. Is Wright’s shifting balance process important in evolution? Evolution 54: 306–317.
Crozier R.H. 1987. Genetic aspects of kin recognition: Concepts, models, and synthesis. In Kin recognition in animals (ed. D.J.C. Fletcher and C.D. Michener), pp. 55–73. Wiley, New York.
Darwin C. 1859. On the origin of species by means of natural selection. John Murray, London.
Dawkins R. 1982. The extended phenotype. Oxford University Press, Oxford.
de la Casa-Esperón E. and Sapienza C. 2003. Natural selection and the evolution of genome imprinting. Annu. Rev. Genet. 37: 349–370.
Diggle S.P., Griffin A.S., Campbell G.S., and West S.A. 2007. Cooperation and conflict in quorum sensing bacterial populations. Nature 450: 411–415.
Dugatkin L.A. 2007. Inclusive fitness theory from Darwin to Hamilton. Genetics 176: 1375–1380.
Eickbush T.H. and Furano A.V. 2002. Fruit flies and humans respond differently to retrotransposons. Curr. Opin. Genet. Dev. 12: 669–674.
Eigen M. 1992. Steps towards life. Oxford University Press, Oxford.
Eigen M., Gardiner W., Schuster P., and Winkler-Oswatitsch R. 1981. The origin of genetic information. Sci. Am. 244: 88–92, 96, et passim.
Engels W.R. 1996. P elements in Drosophila. Curr. Top. Microbiol. Immunol. 204: 104–123.
Engels W.R. 1997. Invasions of P elements. Genetics 145: 11–15.
Fisher R.A. 1930. The genetical theory of natural selection. Oxford University Press, Oxford.
Frank S.A. 1997. The Price equation, Fisher’s fundamental theorem, kin selection, and causal analysis. Evolution 51: 1712–1729.
Frank S.A. 1998. Foundations of social evolution. Princeton University Press, Princeton, New Jersey.
Gimble F.S. and Thorner J. 1992. Homing of a DNA endonuclease gene by meiotic gene conversion in Saccharomyces cerevisiae. Nature 357: 301–305.
Godfray H.C.J., Partridge L., and Harvey P.H. 1991. Clutch size. Annu. Rev. Ecol. Syst. 22: 409–430.
Gould S.J. and Eldredge N. 1977. Punctuated equilibria: The tempo and mode of evolution reconsidered. Paleobiology 3: 115–151.
Grafen A. 1979. Hawk-Dove game played between relatives. Anim. Behav. 27: 905–907.
Grafen A. 1982. How not to measure inclusive fitness. Nature 298: 425–426.
Grafen A. 1985. A geometric view of relatedness. Oxf. Surv. Evol. Biol. 2: 28–89.
Gregory T.R. 2001. Coincidence, coevolution, or causation? DNA content, cell size, and the C-value enigma. Biol. Rev. Camb. Philos. Soc. 76: 65–102.
Gregory T.R. 2008. Animal genome size database. http://www.genomesize.com/
Greilhuber J. 1998. Intraspecific variation in genome size: A critical reassesment. Ann. Bot. (Lond.) 82: 27–35.
Griffin A.S. and West S.A. 2002. Kin selection: Fact and fiction. Trends Ecol. Evol. 17: 15–21.
Griffin A.S. and West S.A. 2003. Kin discrimination and the benefit of helping in cooperatively breeding vertebrates. Science 302: 634–636.
Haldane J.B.S. 1949. Disease and evolution. Ricerca Scient. Suppl. 19: 68–76.
Haldane J.B.S. 1955. Population genetics. New Biol. 18: 34–51.
Hamilton W.D. 1963. The evolution of altruistic behaviour. Am. Nat. 97: 354–356.
Hamilton W.D. 1964a. The genetical evolution of social behaviour I. J. Theor. Biol. 7: 1–16.
Hamilton W.D. 1964b. The genetical evolution of social behaviour II. J. Theor. Biol. 7: 17–52.
Hamilton W.D. 1996a. Narrow roads of gene land, Volume 1: Evolution of social behaviour. W.H. Freeman, Oxford.
Hamilton W.D. 1996b. Narrow roads of gene land, Volume 2: Evolution of sex and sexual selection. W.H. Freeman, Oxford.
Hart M.W. and Grosberg R.K. 1999. Kin interactions in a colonial hydrozoan (Hydractinia symbiolongicarpus): Population structure on a mobile landscape. Evolution 53: 793–805.
Hatch F.T., Bodmer A.J., Magrimas J.A., and Moore D.H. 1976. Satellite DNA and cytogenetic evolution. DNA quantity satellite DNA and karyotype variations in kangaroo rats (genus Dipodomys). Chromosoma 58: 155–168.
Herre E.A. 1985. Sex ratio adjustment in fig wasps. Science 228: 896–898.
Hewitt G.M. and East T.M. 1978. Effects of B chromosomes on development in grasshopper embryos. Heredity 41: 347–356.
Hurst G.D.D. and Werren J.H. 2001. The role of selfish genetic elements in eukaryotic evolution. Nat. Rev. Genet. 2: 597–606.
Hurst L.D. and Pomiankowski A. 1998. Sexual selection: The eyes have it. Nature 391: 223–224.
Jaenike J. 2001. Sex chromosome meiotic drive. Annu. Rev. Ecol. Syst. 32: 25–49.
Kiers E.T., Rousseau R.A., West S.A., and Denison R.F. 2003. Host sanctions and the legume–rhizobium mutualism. Nature 425: 78–81.
Kilner R.M., Madden J.R., and Hauber M.E. 2004. Brood parasitic cowbird nestlings use host young to procure resources. Science 305: 877–879.
King G.J. 2002. Through a genome, darkly: Comparative analysis of plant chromosomal DNA. Plant Mol. Biol. 48: 5–20.
Kirkpatrick S., Gelatt C.D., and Vecchi M.P. 1983. Optimization by simulated annealing. Science 220: 671–680.
Koufopanou V., Goddard M.R., and Burt A. 2002. Adaptation for horizontal transfer in a homing endonuclease. Mol. Biol. Evol. 19: 239–246.
Lander E.S., Linton L.M., Birren B., Nusbaum C., Zody M.C., et al.; International Human Genome Sequencing Consortium. 2001. Initial sequencing and analysis of the human genome. Nature 409: 860–921.
Lehmann L., Keller L., West S., and Roze D. 2007. Group selection and kin selection: Two concepts but one process. Proc. Natl. Acad. Sci. 104: 6736–6739.
Leigh E.G. 1983. When does the good of the group override the advantage of the individual? Proc. Natl. Acad. Sci. 80: 2985–2989.
Lewontin R.C. 1962. Interdeme selection controlling a polymorphism in the housemouse. Am. Nat. 96: 65–78.
Lynch M. and Conery J.S. 2003. The origins of genome complexity. Science 302: 1401–1404.
Lyon M.F. 2003. Transmission ratio distortion in mice. Annu. Rev. Genet. 37: 393–408.
Lyttle T.W. 1979. Experimental population genetics of meiotic drive systems. II. Accumulation of genetic modifiers of Segregation Distorter (SD) in laboratory populations. Genetics 91: 339–357.
Mallet J.L.B. and Joron M. 1999. Evolution of diversity in warning color and mimicry: Polymorphisms, shifting balance, and speciation. Annu. Rev. Ecol. Syst. 30: 201–233.
Maschwitz U. and Maschwitz E. 1974. Platzende Arbeiterinen: Eine Neue Art der Feindabwehr bei sozialen Hautflüglern. Oecologia 14: 289–294.
Maside X., Assimacopoulos S., and Charlesworth B. 2000. Rates of movement of transposable elements on the second chromosome of D. melanogaster. Genet. Res. 75: 275–284.
Maynard Smith J. 1976. Group selection. Q. Rev. Biol. 51: 277–283.
Maynard Smith J. and Szathmáry E. 1995. The major transitions in evolution. W.H. Freeman, Oxford.
Montchamp-Moreau C., Ginhoux V., and Atlan A. 2001. The Y chromosomes of Drosophila simulans are highly polymorphic for their ability to suppress sex-ratio drive. Evolution 55: 728–737.
Nee S. and Maynard Smith J. 1990. The evolutionary biology of molecular parasites. Parasitology (suppl.) 100: S5–S18.
Nobrega M.A., Zhu Y., Plajzer-Frick I., Afzal V., and Rubin E.M. 2004. Megabase deletions of gene deserts result in viable mice. Nature 431: 988–993.
Nur U., Werren J.H., Eickbush D.G., Burke W.D., and Eickbush T.H. 1988. A “selfish” B chromosome that enhances its transmission by eliminating the paternal genome. Science 240: 512–514.
Oster G.F. and Wilson E.O. 1979. Caste and ecology in the social insects. Princeton University Press, New Jersey.
Palopoli M.F. and Wu C.I. 1996. Rapid evolution of a coadapted gene complex: Evidence from the Segregation Distorter (SD) system of meiotic drive in Drosophila melanogaster. Genetics 143: 1675–1688.
Pardue M.L. and Debarshye P.G. 2003. Retrotransposons provide an evolutionarily robust non-telomerase mechanism to maintain telomeres. Annu. Rev. Genet. 37: 485–511.
Parker G.A. and MacNair M.R. 1978a. Models of parent-offspring conflict. I. Monogamy. Anim. Behav. 26: 97–110.
Parker G.A. and MacNair M.R. 1978b. Models of parent-offspring conflict. II. Polygamy. Anim. Behav. 26: 111–122.
Partridge L. and Hurst L.D. 1998. Sex and conflict. Science 281: 2003–2008.
Pellmyr O. and Huth C.J. 1994. Evolutionary stability of mutualism between yuccas and yucca moth. Nature 372: 257–260.
Pellmyr O. and Leebensmack J. 1999. Forty million years of mutualism: Evidence for Eocene origin of the yucca-yucca moth association. Proc. Natl. Acad. Sci. 96: 9178–9183.
Pennisi E. 2007. Natural selection, not chance, paints the desert landscape. Science 318: 376.
Pfeiffer T. and Bonhoeffer S. 2003. An evolutionary scenario for the transition to undifferentiated multicellularity. Proc. Natl. Acad. Sci. 100: 1095–1098.
Pfennig D.W. 1999. Cannibalistic tadpoles that pose the greatest threat to kin are most likely to discriminate kin. Proc. R. Soc. Lond. B 266: 57–61.
Plasterk R.H.A. 2002. RNA silencing: The genome’s immune system. Science 296: 1263–1265.
Poggio L., Rosato M., Chiavarino A.M., and Naranjo C.A. 1998. Genome size and environmental correlations in maize (Zea mays ssp. mays, Poaceae). Ann. Bot. (Lond.) 82 (A): 107–115.
Price G.R. 1970. Selection and covariance. Nature 227: 520–521.
Provine W. 1986a. Evolution: Collected papers of S. Wright. University of Chicago Press, Chicago.
Provine W. 1986b. Sewall Wright and evolutionary biology. University of Chicago Press, Chicago.
Queller D.C., Ponte E., Bozzaro S., and Strassmann J.E. 2003. Single-gene Greenbeard effects in the social amoeba Dictyostelium discoideum. Science 274: 993–995.
Ratnieks F.L.W. 1988. Reproductive harmony via mutual policing by workers in eusocial Hymenoptera. Am. Nat. 132: 217–236.
Ratnieks F.L.W. and Wenseleers T. 2005. Policing insect societies. Science 307: 54–56.
Roth G., Nishikawa K.C., and Wake D.B. 1997. Genome size, secondary simplification, and the evolution of the brain in salamanders. Brain Behav. Evol. 50: 50–59.
Russell A.F. and Hatchwell B.J. 2001. Experimental evidence for kin-biased helping in a cooperatively breeding vertebrate. Proc. R. Soc. Lond. B 268: 2169–2174.
Sandoz K.M., Mitzimberg S.M., and Schuster M. 2007. Social cheating in Pseudomonas aeruginosa quorum sensing. Proc. Natl. Acad. Sci. 104: 15876–15881.
San Miguel P. and Bennetzen J.L. 1998. Evidence that a recent increase in maize genome size was caused by the massive amplification of intergene retrotransposons. Ann. Bot. (Lond.) 82 (A): 37–44.
San Miguel P., Gaut B.S., Tikhonov A.P., Nakajima Y., and Bennetzen J.L. 2000. The paleontology of intergene retrotransposons of maize. Nat. Genet. 20: 43–45.
Schemske D.W. and Bierzychudek P. 2001. Evolution of flower color in the desert annual Linanthus parryae: Wright revised. Evolution 55: 1269–1282.
Schemske D.W. and Bierzychudek P. 2007. Spatial differentiation for flower color in the desert annual Linanthus parryae: Was Wright right? Evolution 61: 2526–2543.
Selker E.U. 2002. Repeat-induced gene silencing in fungi. Adv. Genet. 46: 439–450.
Servedio M.R. 2000. The effects of predator learning, forgetting, and recognition errors on the evolution of warning coloration. Evolution 54: 751–763.
Sinervo B. and Clobert J. 2003. Morphs, dispersal behavior, genetic similarity, and the evolution of cooperation. Science 300: 1949–1951.
Sleutels F. and Barlow D.P. 2002. The origins of genomic imprinting in mammals. Adv. Genet. 46: 120–163.
Smith N.G.C. and Hurst L.D. 1998. Molecular evolution of an imprinted gene: Repeatability of patterns of evolution within the mammalian insulin-like growth factor type II receptor. Genetics 150: 823–833.
Spencer H.G. 2000. Population genetics and evolution of genomic imprinting. Annu. Rev. Genet. 34: 457–478.
Spencer H.G., Clark A.G., and Feldman M.W. 1999. Genetic conflicts and the evolutionary origin of genomic imprinting. Trends Ecol. Evol. 14: 197–201.
Stadler T. and Delph L.F. 2002. Ancient mitochondrial haplotypes and evidence for intragenic recombination in a gynodioecious plant. Proc. Natl. Acad. Sci. 99: 11730–11735.
Stanley S. 1975. A theory of speciation above the species level. Proc. Natl. Acad. Sci. 72: 646–650.
Stanley S. 1979. Macroevolution. W.H. Freeman, San Francisco.
Thompson J.D., Manicacci D., and Tarayre M. 1998. Thirty-five years of thyme: A tale of two polymorphisms. Bioscience 48: 805–815.
Turelli M. and Hoffmann A.A. 1995. Cytoplasmic incompatibility in Drosophila simulans: Dynamics and parameter estimates from natural populations. Genetics 140: 1319–1338.
Turelli M., Hoffmann A.A., and McKechnie S.W. 1992. Dynamics of cytoplasmic incompatibility and mtDNA variation in natural Drosophila simulans populations. Genetics 132: 713–723.
Van Damme J.M.M., Hundscheid M.P.J., Ivanovic S., and Koelewijn H.P. 2004. Multiple CMS-restorer gene polymorphism in gynodioecious Plantago coronopus. Heredity 93: 175–181.
Watson J.D., Baker T.A., Bell S.P., Gann A., Levine M., and Losick R. 2008. Molecular biology of the gene, 6th edition. Pearson/Benjamin Cummings, San Francisco, California (co-published with Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York).
Weeks A.R., Reynolds K.T., Hoffmann A.A., and Mann H. 2002. Wolbachia dynamics and host effects: What has (and has not) been demonstrated? Trends Ecol. Evol. 17: 257–262.
Welch D.M. and Meselson M. 2000. Evidence for the evolution of bdelloid rotifers without sexual reproduction or genetic exchange. Science 288: 1211–1215.
West S.A., Griffin A.S., and Gardner A. 2007. Social semantics: Altruism, cooperation, mutualism, strong reciprocity and group selection. J. Evol. Biol. 20: 415–432.
West S.A., Griffin A.S., and Gardner A. 2008. Social semantics: How useful has group selection been? J. Evol. Biol. 21: 374–385.
West S.A., Murray M.G., Machado C.A., Griffin A.S., and Herre E.A. 2001. Testing Hamilton’s rule with competition between relatives. Nature 409: 510–513.
Wilkinson G.S., Presgraves D.C., and Crymes L. 1998. Male eye span in stalk-eyed flies indicates genetic quality by meiotic drive suppression. Nature 391: 276–279.
Williams G.C. 1992. Natural selection: Domains, levels and challenges. Oxford University Press, Oxford.
Wilson D.S. 1983. The group selection controversy: History and current status. Annu. Rev. Ecol. Syst. 14: 159–188.
Wilson D.S. 2008. Social semantics: Towards a genuine pluralism in the study of social behaviour. J. Evol. Biol. 21: 368–373.
Wilson D.S. and Wilson E.O. 2007a. Rethinking the theoretical foundation of sociobiology. Q. Rev. Biol. 82: 327–348.
Wilson D.S. and Wilson E.O. 2007b. Survival of the selfless. New Scientist 196: 42–46.
Wilson E.O. 1971. The insect societies. Harvard University Press, Cambridge, Massachusetts.
Wilson E.O. 1975. Sociobiology: The new synthesis. Harvard University Press, Cambridge, Massachusetts.
Wright S. 1931. Evolution in Mendelian populations. Genetics 16: 97–159.
Wright S. 1932. The roles of mutation, inbreeding, crossbreeding and selection in evolution. Proc. Sixth Int. Congr. Genet. 1: 356–366.
|



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}